For each new batch of cell extract and T7 RNA polymerase, it is recommended to perform a basic screening of both Mg2+ and K+ concentrations to ensure the optimal performance of the CFPS system. The fluorescence of superfolder GFP can serve as an indicator of the overall yield of the CFPS system under varying conditions, as illustrated in Figure 1A,B. Additionally, a parallel yield comparison of the CFPS system across different compartments is shown in Figure 1C, demonstrating consistent performance in encapsulated CFPS systems, albeit with a slight decrease in GUVs, potentially due to the influence of sucrose46. Furthermore, CFPS reactions were successfully encapsulated in droplets (Figure 2), GUVs (Figure 3), and atop SLBs (Figure 4).
Figure 2A highlights the formation of emulsion droplets, evident when the oil solution became turbulent. The expression of green/red fluorescent proteins (GFP and mCherry) was captured using a confocal laser scanning microscopy. Figure 3 showcased the success of the encapsulation of CFPS reaction within GUVs through the simple emulsion transfer process. The red channel indicated the labeled lipids; the green channel showed the expression of superfolder GFP; and the yellow channel represented an inert fluorescent dye. Figure 4A shows the SLB chamber where the CFPS reaction could be deposited. Within this chamber, an SLB was formed on top of the glass coverslip, with the CFPS reaction mixture placed atop the preformed SLB, enabling imaging via a confocal laser scanning microscopy. As indicated in Figure 4B, SLB was visualized by labeled lipids, shown in green, while the expressed small GTPase Cdc42 fused with mCherry was shown in magenta. The in-situ expression of mCherry-Cdc42 was observable, along with the reversible membrane targeting process, under the microscope. However, the CFPS system utilized was supplemented with prenylation machinery, as detailed in a recent publication47.

Figure 1: Optimization and comparison of the CFPS system. Screening of (A) Mg2+ concentration and (B) K+ concentrations for the CFPS system in bulk. (C) A bar chart showing protein expression yields in different compartments. The fluorescence of superfolder GFP was used to determine the yield of the CFPS system, measured by a plate reader or a confocal laser scanning microscope. Abbreviations: CFPS = cell-free protein synthesis; GFP = green fluorescent protein; sfGFP = superfolder GFP; GUV = giant unilamellar vesicle; SLB = supported lipid bilayer. Please click here to view a larger version of this figure.

Figure 2: Cell-free protein synthesis in droplets. (A) A photo of encapsulated CFPS reactions in surfactant-stabilized water-in-oil droplets. (B) Brightfield and (C,D) fluorescence images of CFPS of (C) mCherry and (D) superfolder GFP encapsulated in droplets visualized by confocal laser scanning microscopy. Superfolder GFP is indicated in green; mCherry is indicated in magenta. Scale bars = 20 µm. Abbreviations: CFPS = cell-free protein synthesis; GFP = green fluorescent protein. Please click here to view a larger version of this figure.

Figure 3: Cell-free protein synthesis in GUVs. (A) A photo of encapsulated CFPS reactions in GUVs. (B–D) Fluorescence image of encapsulated CFPS within POPC GUVs, expressing superfolder GFP. (B) The lipid bilayer was labeled with 0.05% labeled Atto647N-DOPE lipids, shown in red, (C) an inert dye shown in yellow, and (D) superfolder GFP was indicated in green. Scale bars = 20 µm. Abbreviations: CFPS = cell-free protein synthesis; GFP = green fluorescent protein; GUV = giant unilamellar vesicle; POPC = 1-palmitoyl-2-oleoyl-glycerol-3-phosphocholine; DOPE = 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine. Please click here to view a larger version of this figure.

Figure 4: Cell-free protein synthesis of small GTPase protein-Cdc42 on top of SLBs. (A) Schematic illustration of the reconstitution of Cdc42's membrane targeting and RhoGDI-dependent membrane extraction on SLBs, as visualized by confocal laser scanning microscopy. The prenylation machinery was implemented via a prenlytransferase-enriched extract, which is detailed described elsewhere.47. (B) An orthogonal view of the pre- and postprenylation of the reaction mixture on top of SLB. (C,D) Time series of mCherry-Cdc42 intensity on the membrane (pink) and in solution (black) during (C) membrane targeting and (D) membrane extraction. Scale bars = 10 µm. (A–D) were adapted from Kai et al.47. Please click here to view a larger version of this figure.
Table 1: List of buffers. NOTE: Add β-mercaptoethanol and DTT just before use. Please click here to download this Table.
Table 2: Pipetting scheme for Mg2+ concentration screening. Please click here to download this Table.
Table 3: Pipetting scheme for K+ concentration screening. Please click here to download this Table.
Table 4: Inner-solution composition. Please click here to download this Table.
Table 5: Outer solution composition. Please click here to download this Table.
Table 6: Preinner solution composition. Please click here to download this Table.
| 1,2-dioleoyl-sn-glycero-3-phosphocholine(DOPC) | Avanti | 850375P | |
| 1,2-dioleoyl-sn-glycero-3-phospho-L-serine (sodium salt)(DOPS) | Avanti | 840035P | |
| 1,4 dithiothreitol (DTT) | Sigma-Aldrich | 1.11474 | |
| 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC) | Avanti | 850457P | |
| 3,5-cyclic AMP (cAMP) | Sigma-Aldrich | A9501 | |
| 50 mL tubes | Eppendorf | Eppendorf Tubes BioBased | |
| 50% hydrogen peroxide | Sigma-Aldrich | 516813 | |
| Acetate | Sigma-Aldrich | A6283 | |
| Agar powder | Sigma-Aldrich | 05040 | |
| Alanin | Sigma-Aldrich | A4349 | |
| Amicon Stirred Cells | MerckMillipore | UFSC05001 | |
| Ammonium acetate (NH4OAc) | Sigma-Aldrich | A7262 | |
| Arginin | Sigma-Aldrich | A4474 | |
| Asparagin | Sigma-Aldrich | A0884 | |
| Aspartat | Sigma-Aldrich | A5474 | |
| ATP | Roche | 11140965001 | |
| Atto 488 DOPE | Sigma-Aldrich | 67335 | |
| Atto 647N DOPE | Sigma-Aldrich | 42247 | |
| Baffled Erlenmeyer flask | Shuniu | 250 mL, 1000mL | |
| Bovine Serum Albumin(BSA) | Roche | 10711454001 | |
| Centrifugetube | Eppendorf | Eppendorf Tubes 3810X | |
| Centrifugetube rack | Eppendorf | 0030119819 | |
| Chemiluminescence and epifluorescence imaging system | Uvitec | Alliance Q9 Advanced | |
| Chloroform | Sigma-Aldrich | 288306 | |
| Confocal Laser Scanning Microscopy (LSM) | ZEISS | LSM 780 | |
| Countess Cell Counting Chamber Slides | Thermo Fisher Scientific | C10283 | |
| Coverslip | Thermo Scientific | Menzel BB02400500A113MNZ0 | |
| creatine kinase (CK) | Roche | 10127566001 | |
| Creatine phosphate (CP) | Sigma-Aldrich | 10621714001 | |
| Culture dish | Huanqiu | 90 mm | |
| Cystein | Sigma-Aldrich | C5360 | |
| Cytidine 5'-triphosphate disodium salt (CTP) | aladdin | C101487 | |
| Dialysis membrane | Spectrum | Standard RC Tubing MWCO: 12-14 kD | |
| E.Z.N.A. Cycle Pure Kit | Omega Bio-Tek | D6492-01 | |
| Electro-Heating Standing-Temperature Cultivator | Yiheng instrument | DHP-9602 | |
| Ethylenediaminetetraacetic acid(EDTA) | Biosharp | 1100027 | |
| Fluorescent plate reader | BioTek | Synergy 2 | |
| Fluorinated oil | Suzhou CChip scientific instrument | 2%HFE7500 | |
| Folinic acid | Sigma-Aldrich | 47612 | |
| French Press | G.Heinemann | HTU-DIGI-Press | |
| Glucose | Sigma-Aldrich | G7021 | |
| Glutamat | Sigma-Aldrich | G5667 | |
| Glutamin | Sigma-Aldrich | G5792 | |
| Glycerol | Sigma-Aldrich | G5516 | |
| Glycin | Sigma-Aldrich | G7126 | |
| Guanosine 5'-triphosphate sodium salt hydrate(GTP) | Roche | 10106399001 | |
| HEPES | Sigma-Aldrich | H3375 | |
| HiPrep Q FF 16/10 | Cytiva | 28936543 | |
| Histidin | Sigma-Aldrich | H6034 | |
| Isoleucin | Sigma-Aldrich | I5281 | |
| Isopropyl-β-D-thiogalactopyranoside (IPTG) | Sigma-Aldrich | I5502 | |
| K2HPO4 | Sigma-Aldrich | P8281 | |
| KH2PO4 | Sigma-Aldrich | P5655 | |
| Leucin | Sigma-Aldrich | L6914 | |
| Lysin | Sigma-Aldrich | L5501 | |
| Magnesium acetate tetrahydrate (Mg(OAc)2 ) | Sigma-Aldrich | M5661 | |
| Magnesium chloride(MgCl2) | Sigma-Aldrich | M2670 | |
| Methionin | Sigma-Aldrich | M8439 | |
| Microcentrifuge | Eppendorf | 5424 R | |
| Mineral oil | Sigma-Aldrich | M5904 | |
| Mini-PROTEAN Tetra Cell Systems | Bio-Rad | 1645050 | |
| Multipurpose Centrifuge | Eppendorf | 5810 R | |
| NaN3 | Sigma-Aldrich | S2002 | |
| Nucleic Acid & Protein UV-Assay Measurements | IMPLEN | NanoPhotometer N60 | |
| NucleoBond Xtra Maxi kit for transfection-grade plasmid DNA | MACHEREY-NAGEL | 740414.5 | |
| Nunc-Immuno MicroWell 96 well polystyrene plates | Sigma-Aldrich | P8616 | |
| PCR Thermal Cycler | Eppendorf | Mastercycler nexus | |
| Peptone | Sigma-Aldrich | 83059 | |
| Phenylalanin | Sigma-Aldrich | P8740 | |
| Phosphoenolpyruvat (PEP) | GLPBIO | GC44635 | |
| PMSF | Sigma-Aldrich | PMSF-RO | |
| Polyethylene glycol 8000 (PEG 8000) | Sigma-Aldrich | 89510 | |
| Potassium Acetate(KOAc) | Sigma-Aldrich | P5708 | |
| Potassium chloride(KCl) | Sigma-Aldrich | P9541 | |
| Potassium glutamate (K-glutamate) | Sigma-Aldrich | G1501 | |
| Potassium hydroxide(KOH) | Sigma-Aldrich | 221473 | |
| Prolin | Sigma-Aldrich | P8865 | |
| Pyruvate kinase (PK) | Sigma-Aldrich | P9136 | |
| Serin | Sigma-Aldrich | S4311 | |
| Shaker | Zhichushakers | ZQZY-AF8 | |
| Sodium chloride(NaCl) | Sigma-Aldrich | S5886 | |
| Sodium hydroxide(NaOH) | Sigma-Aldrich | S5881 | |
| Sucrose | aladdin | S112226 | |
| Sulfuric acid | Sigma-Aldrich | 339741 | |
| Syringe Filters | Jinteng | 0.45 μm | |
| Test tube | Shuniu | 20 mL | |
| TGX FastCast Acrylamide Kit, 12% | Bio-Rad | #1610175 | |
| ThermoMixer | Eppendorf | ThermoMixer C | |
| Threonin | Sigma-Aldrich | T8441 | |
| Tris base | Sigma-Aldrich | V900483 | |
| tRNA | Roche | 10109550001 | |
| Tryptone | Sigma-Aldrich | T7293 | |
| Tryptophan | Sigma-Aldrich | T8941 | |
| Tyrosin | Sigma-Aldrich | T8566 | |
| UTP Trisodium salt (UTP) | aladdin | U100365 | |
| Vacuum Pump with Circulated Water System | Zhengzhou Greatwall Scientific Industrial and Trade Co.Ltd | SHB-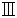 |
|
| Valin | Sigma-Aldrich | V4638 | |
| Vortex Mixers | Kylin-Bell | Vortex QL-861 | |
| Water purification system | MerckMillipore | Direct ultrapure water (Type 1) | |
| Yeast extract | Sigma-Aldrich | 70161 | |
| β-mercaptoethanol | Sigma-Aldrich | 444203 |
