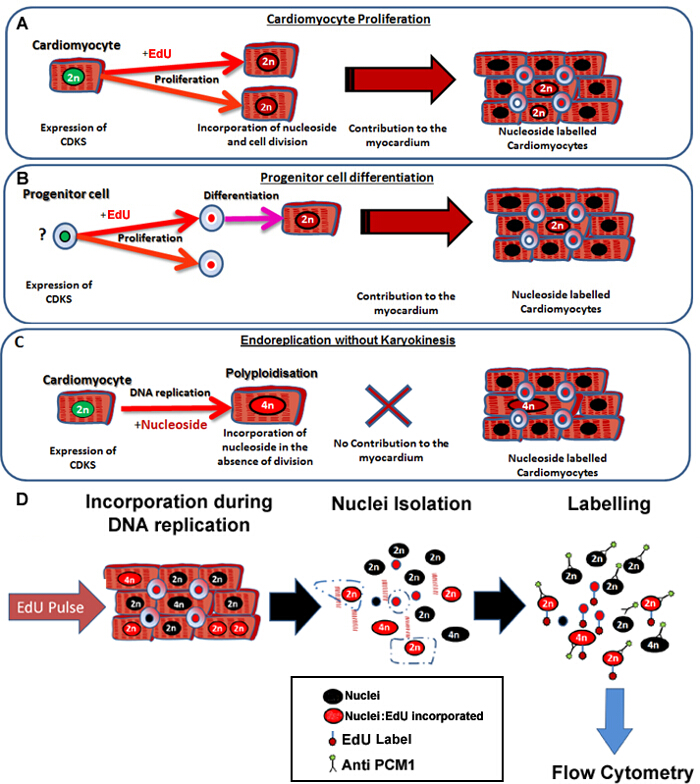
el trabajo con animales fue autorizado y aprobado por el comité de revisión ética de la Universidad de Newcastle. Todos los procedimientos con animales se realizaron conforme a las líneas de la Directiva 2010/63 / UE del Parlamento Europeo sobre la protección de los animales utilizados para fines científicos. 1. Administración EdU Disolver EdU en solución salina estéril (0,9% w / v) a una concentración final de 12,5 mg / ml. Calentar la solución a 40 ° C y agitar para disolver completamente. Almacenar suficiente EDU / solución salina a 4 ° C para inyectar todos los ratones en el estudio lo que permite 6 inyecciones diarias. Nota: Por lo general se requieren más de 6 ratones para cada grupo experimental. Sin embargo, los cálculos de potencia se deben realizar para estudios independientes. Pesar ratones individuales. Calcular el volumen necesario para administrar 100 mg / g de solución / solución salina EdU a cada ratón (8 l de solución madre EdU por g). Dibujar el volum apropiadae del EDU / solución salina en jeringa de insulina con una aguja de 25 G. Realizar intraperitoneal (ip) durante la manipulación de los ratones usando técnicas del Ministerio del Interior aprobados. Coloque el ratón en la tapa de la jaula. Tire suavemente de nuevo en la base de la cola y sujete firmemente el ratón por la piel. Exponer el abdomen para inyección Manteniendo el ratón con el dedo índice y el pulgar cerca de la base de la cabeza e inclinando la cabeza del animal hacia atrás hacia el piso. Limpiar el abdomen con una solución de alcohol 70%. Localiza la línea media del animal y en el cuadrante inferior derecho. Se inyecta la solución EDU / solución salina en el animal en el cuadrante inferior derecho. Inyectar ratones con solución / solución salina EdU y registrar la hora del día. Repetir los pasos 1.2 a 1.4 en el mismo momento del día, durante 6 días consecutivos. Nota: A medida que se requiere un control negativo EdU para ajustar la compuerta para el análisis de citometría de flujo, inyectar una edad adicional, el género y experimentalmente que coincidaed ratón con un volumen equivalente de solución salina sin edu. Esto proporcionará el control requerido (ningún control EDU). Este control debe ser incluido para cada estudio para permitir variaciones en los lotes de reactivos y citómetro configurado. 2. Los corazones de cosecha A las 24 h después de la inyección EdU 6º, sacrificar a los ratones por dislocación cervical (o un horario alternativo 1 de Ministerio del Interior aprobó el método). Lugar de los animales sacrificados en una posición supina y hacer incisión en la piel desde el centro del abdomen hasta el diafragma con tijeras quirúrgicas. Cortar el diafragma, y sosteniendo el esternón lejos de la cavidad del cuerpo, corte bilateralmente para exponer el corazón. Levantar ligeramente el corazón y cortar los principales vasos sanguíneos del tracto de salida para diseccionar el corazón fuera de la cavidad torácica. Inmediatamente colocar el corazón en cada una de 15 ml tubo de centrífuga individual que contiene 10 ml de PBS enfriado a 4 ° C. Transferir los corazones a SEParate 10 cm placas de Petri, utilizando un escalpelo cortar cada corazón en dos en una orientación sagital y lavar el corazón apretando suavemente con un par de fórceps. A continuación, reemplace el PBS para eliminar la mayor cantidad de sangre posible. Repita este paso dos veces. Coloque ambas partes de cada corazón en el mismo tubo de microcentrífuga de 1,5 ml individual. Continúe con el paso 3 o, alternativamente, almacenar las muestras -80 ° C como se detalla en los pasos 2.9 y 2.10. Usando una aguja 25 G, hacer un pequeño agujero en la tapa del tubo de microcentrífuga para evitar que la tapa de apertura debido a la expansión de los gases cuando se procesa el corazón. Etiqueta de cada tubo de microcentrífuga y colocar en un contenedor de nitrógeno líquido para congelar. Etiquetado 3. Los núcleos de disociación de los cardiomiocitos y Núcleos Nota: Este etiquetado de disociación y cardiomiocitos núcleos núcleos es una adaptación de la de Bergmann et al 11 Realice este procedimiento en todas las muestras inyectadas.con Edu y solución salina inyectada (sin control de EDU) control negativo. Ver Tabla 1 para la receta de soluciones que se requieren para llevar a cabo estos pasos. Para cada corazón para ser analizada, colocar 36 ml de 1% de BSA / PBS solución en un tubo de ultracentrífuga, se incuba durante 30 minutos, desechar la solución y luego dejar secar al aire. Nota: Este tubo se utiliza en el paso 3.6. Coloque los corazones congelados individuales sobre una placa de 10 mm en el hielo y carne picada con un bisturí que permite a los corazones se descongelan durante el proceso de picar carne. Coloque corazones picado en 15 ml de Tampón de Lisis Celular en un tubo de centrífuga de 50 ml y homogeneizar con un homogeneizador de sonda para 15 segundos a 25.000 rpm a temperatura ambiente. Añadir otros 15 ml de Tampón de Lisis Celular al tubo de centrífuga y transferir a un 40 ml Dounce con una mano de mortero grande despacho. Realizar 10 golpes con la mano del mortero y filtrar a través de 100 micras a continuación colador 40 micras de células en un tubo de centrífuga de 50 ml. Se centrifuga durante 10 minutos unat 700 xga 4 ° C y separar el sobrenadante. Resuspender el sedimento nuclear en 25 ml de sacarosa solución de gradiente y la capa de los núcleos celulares solución en la parte superior de 10 ml de solución de gradiente de sacarosa fresco en el tubo de ultracentrífuga preparado en 3.1 que contienen. Centrifugar a 13.000 xg durante 1 hora a 4 ° C usando un rotor de tubo de oscilación hacia fuera. Tras la centrifugación, desechar el sobrenadante y resuspender el botón núcleos en 1.300 l de tampón de almacenamiento de núcleos en un tubo de 1.5 ml y etiqueta PCM1. Eliminar 300 l de la suspensión a un nuevo tubo de microcentrífuga y añadir 700 l de tampón de almacenamiento núcleos fresco. Etiquetar este tubo como control ISO. Añadir anti-PCM1 a una concentración final de 8 mg / ml al tubo de microcentrífuga etiquetado PCM1 y añadir el anticuerpo de control de isotipo IgG de conejo a una concentración de 8 mg / ml al tubo de microcentrífuga etiquetado de control ISO. Incubar toda la noche a 4 ° C. Tras incubación, centrifugar a 700 xg durante 10 min, descartar el sobrenadante, sustituir con 1 ml de tampón de almacenamiento núcleos fresco, resuspender y centrifugar de nuevo durante 10 min a 700 x g. Descartar el sobrenadante y reemplazar con 1 ml de tampón de almacenamiento núcleos fresco. Añadir 1 l de F (ab ') 2 Fragmento de IgG de cabra anti-conejo (H + L) de anticuerpos (FITC o tinte verde fluorescente equivalente) para alcanzar una concentración final de 2 mg / ml. Nota: El uso de un F (ab ') 2 fragmento de anticuerpo disminuye el potencial para el etiquetado no específica de las células inmunes incluyendo macrófagos y células B. tubos de envoltura en papel de aluminio para protegerlo de la luz y se incuban durante 1 hora a 4 ° C. 4. Detección de EdU Incorporación Diluir 10 veces más concentrada EdU tampón de reacción 1:10 con agua desionizada. Se centrifuga la PCM-1 e ISO suspensiones núcleos de control etiquetada a 700 xg durante 10 min, descartar el sobrenadante y resuspender el botón núcleos en 1 ml de1% de BSA / PBS solución. Lavar el sedimento de núcleos dos veces. Después del lavado final, descartar el sobrenadante y reemplazar con 100 l de EdU fijador. Envuelva los tubos en papel de aluminio para protegerlo de la luz y se incuba a temperatura ambiente durante 15 min. Lavar las muestras dos veces con 1 ml de 1% BSA / PBS solución, centrifugar a 700 xg y se incuba con una solución de permeabilización a base de saponina 1x a temperatura ambiente durante 15 min. Preparar 1x cóctel etiquetado EdU (Tabla 2). Se requerirá un mínimo de 2 reacciones de incluir el control edu. Añadir 500 l de 1x cóctel EdU etiquetado directamente a cada muestra que ya contiene los núcleos en 100 l de solución de permeabilización a base de saponina 1X y se incuba a temperatura ambiente protegido de la luz durante 30 min. Centrifugar a 700 xg y resuspender en 1 ml de 1% BSA / PBS y repetir este paso dos veces más. Se centrifuga a 700 xg, desechar el sobrenadante y reemplazar con 400 l de solución de tinción de ADN (véase la Tabla de Materiales). Nombre del reactivo 1. Tampón de Lisis Celular 0,32 M de sacarosa 10 mM Tris-HCl (pH = 8) 5 mM CaCl 2 acetato de magnesio 5 mM EDTA 2,0 mM EGTA 0,5 mM DTT 1 mM Solución Gradiente 2.Sucrose 2,1 M de sacarosa 10 mM Tris-HCl (pH = 8) acetato de magnesio 5 mM DTT 1 mM 3. tampón de almacenamiento núcleos (NSB) 0,44 M de sacarosa 10 mM Tris-HCl (pH = 7,2) KCl 70 mM 10 mM de MgCl 2 espermina 1,5 mM Tabla 1: Núcleos ingredientes de la receta tampón de aislamiento para todos los tampones usados en la sección de protocolo 3 (disociación núcleos y etiquetado núcleos de cardiomiocitos).. 5. Análisis de citometría de flujo Nota: Realizar la citometría de flujo en núcleos aislados como se describió previamente 7,11,15. En primer lugar, crear un gráfico que describe dispersión frontal (FSC) y dispersión lateral (SSC) para permitir la discriminación de los núcleos de los desechos (Figura 2A). Crear un plan para discriminar los núcleos individuales de los agregados (Figura 2B) mediante el trazado de 4 ', 6-diamino-2-fenilindol)) área (DAPI (3-405 / 450/50-A) respecto a la altura (33-405 / 450 / 50-H). garantizar that el 3-405 / 450/50-A de la señal se recoge en la escala lineal para permitir la correcta determinación del contenido de ADN. Crear un gráfico que describe 488/535 / 30-A (PCM-1) frente al SSC y mostrar sólo los eventos de la puerta singlete creada en 5.2 con el fin de evaluar PCM-1 de expresión en la población singlete. Ejecutar la muestra de control de isotipo a fin de establecer un PCM-1 puerta positivo (Figura 2C). Ejecutar una pequeña cantidad de PCM1 etiquetado de ejemplo para comprobar la PCM1 expresar la posición de la población y gating. Crear un gráfico que describe SSC-A frente a 405/450 / 50-A (para detectar DAPI). En esta gráfica, pantalla sólo singlete Pcm1 núcleos que expresan, usando las puertas creadas en 5.3. Utilice esta parcela para crear una puerta jerárquico adicional que contiene sólo 2 N núcleos (Figura 2D). Crear un gráfico que describe 488/535 / 30-A contra 1-633 / 660/20-A para permitir la identificación de etiquetado EdU de la población que expresa PCM1 cardiomiocitos. Mostrar sólo los núcleos que son singlete, P CM1 + y 2N usando el gating anteriormente. Muestra de corazón correr desde ningún control EdU para configurar la puerta EdU positivo (Figura 2E). Crear la puerta apropiada para las células EdU +. Grabar y cuantificar singlete, 2N, núcleos PCM-1 que expresan las que han incorporado edu. El cálculo de esta población como un porcentaje del total de singlete, PCM1 + núcleos. Esto proporcionará la velocidad de generación de núcleos neo-cardiomiocitos, durante el período de pulso EdU 7 días, como un porcentaje de núcleos totales cardiomiocitos. Nota: Como la unión a ADN DAPI es estequiométrica la intensidad de fluorescencia es proporcional a la cantidad de ADN. Determinar la ploidía en base a la intensidad de la / 450/50-A de la señal 405, como se ha descrito previamente 7. Para evaluar la ploidía dentro de la población total de cardiomiocitos utilizar la trama establecida en 5,5 y un gating estrategia basada en la concentración de ADN 7. /files/ftp_upload/53979/53979fig2.jpg "/> Figura 2:. Ventana de medida para cuantificar la estrategia de cardiomiocitos EdU incorporación y ploidía (A) Los núcleos están discriminado de desechos basados en la dispersión frontal (FSC) y dispersión lateral (SSC). (B) Se crea una puerta lineal y la población se identifica singlete núcleos basados en el marcaje con DAPI y la altura 405/450/50 vs señal de zona. (C) gating fluorescente permite la separación de los núcleos de cardiomiocitos núcleos (PCM-1-positivo) y no cardiomiocitos (PCM-1-negativos) de tejido del corazón. (D) La intensidad de la señal DAPI se utiliza para determinar la concentración de ADN y la ploidía de ese modo los núcleos de la población de los cardiomiocitos PCM1 +. En el ratón> 80% de los núcleos de cardiomiocitos son diploide (2n). (E) gating fluorescente permite la separación de 2 N, núcleos de cardiomiocitos que han incorporado EdU (PCM + / + EDU = 0,9%) a partir de 2 N, cardiomiocitos que no han incorporado EdU (PCM + educación). Todas las etapas se detallan en el protocolo 5.0. Ejemplo muestra de MDX -. / Y ratones en el fondo C57 / BL10 Haga clic aquí para ver una versión más grande de esta figura.