في السنوات الأخيرة كان هناك تراكم الأدلة تتحدى الافتراض بأن القلب هو متباينة عضال، بعد الإنقسامية الجهاز 1،2. ومع ذلك، وتحديد حجم دوران cardiomyocyte وتجديد يبقى تحديا.
وذكرت الصعوبات في تحديد بدقة الجيل cardiomyocyte نادر باستخدام تقنيات المناعى القياسية أيضا 3. وبالإضافة إلى ذلك، فإن مصدر الخلوية من الجيل cardiomyocyte لا يزال غير مؤكد مع الدليل على مساهمات من انتشار cardiomyocyte فضلا عن الخلايا الجذعية التمايز 4-6. ولذلك، فإن استخدام نماذج النسب تتبع التي تتطلب معرفة النمط الظاهري cardiomyocyte السلف مستحيل وتقدير الانتشار في عدد السكان واحد، بما في ذلك العضلية، غير مناسب. وعلاوة على ذلك لcardiomyocyte ديه القدرة على endoreplication دون الحرائك النووية (مما أدى إلى السيارة المتعددة الصبغياتdiomyocyte) أو الحرائك النووية في غياب الانقسام السيتوبلازمي (مما أدى إلى cardiomyocyte binucleated) 7،8. والتقدير الدقيق دوران cardiomyocyte يعتمد على القدرة على التمييز بين هذه الأحداث وجيل صحيح الجدد cardiomyocyte. وهذا يخلق تعقيدات فريدة من نوعها لتكرار الحمض النووي والتعبير عن تحركات تعتمد على السيكلين في العضلية لا تظهر على وجه الحصر انقسام الخلايا الحقيقية 9،10.
للمساعدة في القياس الكمي للجيل الجدد cardiomyocyte، جمعنا بين تقنية أنشئت العزلة نوى، ووضع العلامات المناعي من المواد محيط بالمريكز 1 (PCM1) لتحديد نوى cardiomyocyte كما وصفها برغمان وآخرون 7،11 مع طرق جديدة للالمدى الطويل وضع العلامات الحمض النووي وتحليل الصيغة الصبغية. PCM-1 هو بروتين الجسيم المركزي التي تتراكم على سطح النووي متباينة، myocytes غير ركوب الدراجات. وقد أظهرت دراسات سابقة أن الأجسام المضادة ضدPCM-1 تسمية تحديدا نوى cardiomyocyte 7،11 وكما تم استخدام هذا PCM1 من قبل عدد من جماعات مستقلة لتحديد العضلية 1،12،13. وبالإضافة إلى ذلك، لقد أثبتنا أن PCM1 التعبير خرائط لنوى cardiomyocyte المسمى وراثيا في نموذج لجنة المساواة العرقية تي ان تي المعدلة وراثيا الماوس 14 (الشكل التكميلي 1).
بروتوكول صفها هنا يمكن تحديد دقيق وحساس من جيل cardiomyocyte نوى الجدد في قلب فأر، بغض النظر عن أصول الخلوية (الشكل 1A و B) مع استبعاد في نفس الوقت وضع العلامات نوكليوزيد بسبب polyploidization من التحليل (الشكل 1C و D). على الرغم من أن هذه الطريقة لا يمكن السيطرة على ازدواجية النواة cardiomyocyte، لأنها تتيح الكمي السريع والقوي من نوى الجدد cardiomyocyte وهو مطلوب لتقدير دقيق لدوران cardiomyocyte. علاوة على ذلك،أنه يوفر أداة الفرز السريع لتقييم التغيرات المحتملة في ديناميات الجيل cardiomyocyte.
في حين وصفها الحمض النووي وعادة ما ينطوي 5-برومو-2'-deoxyuridine (BrdU) كما التناظرية ثيميدين، وبروتوكول الموصوفة هنا يستخدم فحص 5-إيثينيل-2'-deoxyuridine (EDU) استنادا لأنه يتطلب خطوات المعالجة أقل لأسرع من خلال-وضع ولا يتطلب تغيير طبيعة الحمض النووي للالمناعية الكشف، مما يجعلها متوافقة مع بروتوكولات المناعية الأخرى، وبالتالي زيادة التطبيقات المصب المحتملة لهذه الطريقة.
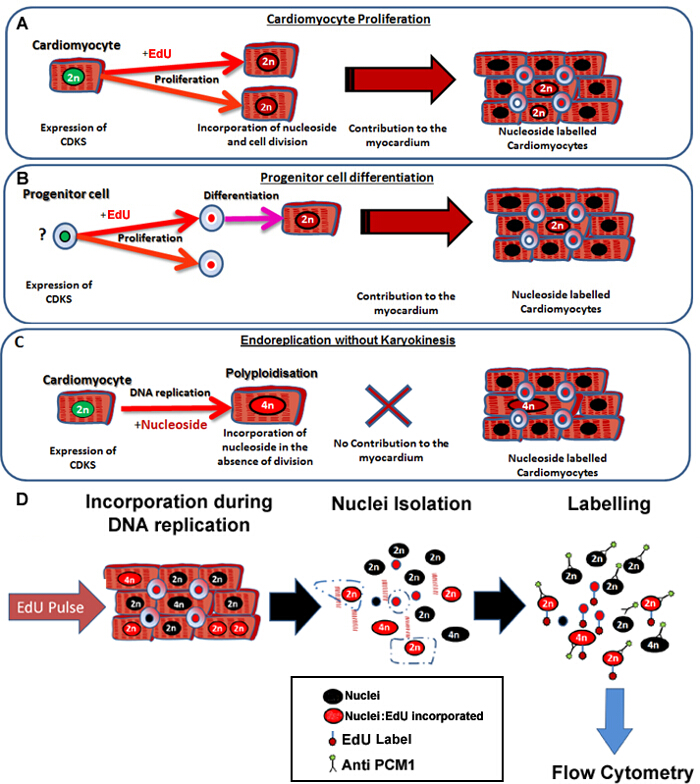
الشكل 1: ينبض المستمر مع ايدو تسميات الجدد العضلية بغض النظر عن الأسلاف الخاصة بهم. أدرج (A) EDU في الحمض النووي العضلية أثناء انقسام الخلية. وانتشار بين السكان cardiomyocyte بالنتيجهتي في زيادة أو استبدال الخلايا العضلية وهو توليف الحمض النووي وبالتالي إنتاجية (يساهم في صيانة الأنسجة وإصلاح). أدرج (ب) EDU في الحمض النووي للخلايا السلف القلب أثناء انقسام الخلية. سيتم الاحتفاظ هذا في الخلية خلال التمايز إلى النسب cardiomyocyte. وهذا تمايز الخلايا الجذعية يؤدي أيضا إلى زيادة في عدد الخلايا العضلية، وبالتالي يساهم في صيانة الأنسجة والإصلاح. (C) العضلية لديها القدرة على الخضوع لتكرار الحمض النووي "غير منتجة" مما أدى إلى زيادة الصيغة الصبغية cardiomyocyte، الذي يرتبط مع تضخم cardiomyocyte وإعادة عضلة القلب، ولكن لا يحل محل العضلية المفقودة. عملية polyploidization تختلف عن ازدواجية النواة كما أنه يؤدي إلى cardiomyocyte مع نواة واحدة والذي يحتوي على أربعة أو أكثر من مجموعات من اثنين من الكروموزومات المتماثلة (> 2N). (D) وبعد نوى ص المستمرulse، يصف هذا البروتوكول العزلة النوى وتحديد نوى cardiomyocyte من التعبير PCM1 للسماح الكمي لكلا الصيغة الصبغية cardiomyocyte وايدو التأسيس. PCM1 التعبير وايدو التأسيس الكشف باستخدام التدفق الخلوي. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
