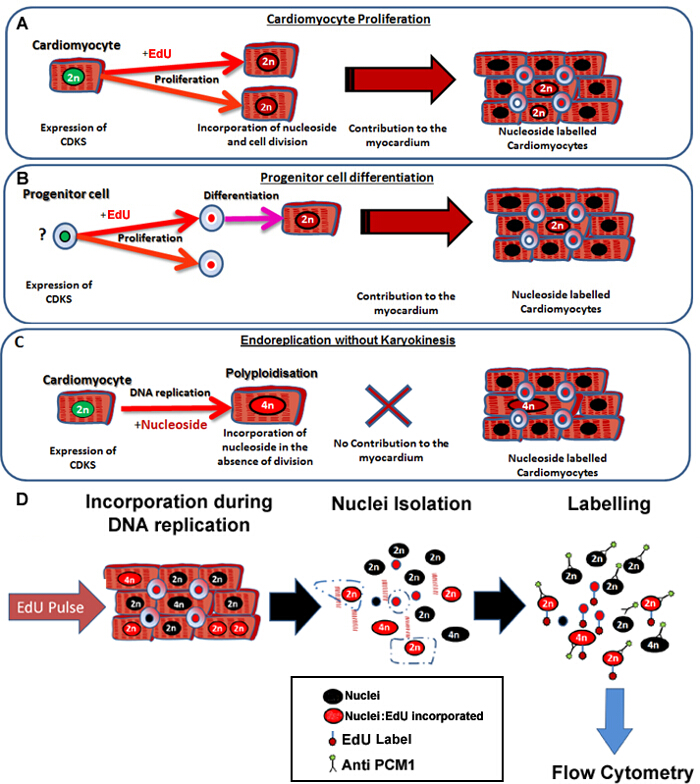
Animal toestemming is verleend en door de Universiteit van Newcastle Ethics Review Board goedgekeurd. Alle dierlijke procedures werden uitgevoerd in overeenstemming met de richtlijnen van Richtlijn 2010/63 / EU van het Europees Parlement over de bescherming van dieren die voor wetenschappelijke doeleinden worden gebruikt. 1. EDU Administration Edu lossen in steriele zoutoplossing (0,9% w / v) bij een eindconcentratie van 12,5 mg / ml. Verwarm de oplossing tot 40 ° C en vortex om volledig op te lossen. WINKEL genoeg edu / zoutoplossing bij 4 ° C om alle muizen te injecteren in de studie waardoor 6 dagelijkse injecties. Opmerking: normaal meer dan 6 muizen worden voor elke experimentele groep. Echter dient vermogen berekeningen worden uitgevoerd voor onafhankelijke studies. Weeg individuele muizen. Bereken het benodigde volume tot 100 ug / g van Edu / zoutoplossing aan elke muis (8 ul van voorraad Edu oplossing per g) beheren. Teken de juiste volume van Edu / zoutoplossing in insulinespuit met een 25 G naald. Voer intraperitoneale (ip) injecties, terwijl de behandeling van de muizen met behulp van goedgekeurde Thuiskantoor technieken. Plaats een muis in de kooi deksel. Trek terug op de basis van de staart en stevig vastpakken van de muis door het nekvel. Expose de buik voor injectie door het vasthouden van de muis met de wijsvinger en duim dichtbij de basis van de kop en storten kop van het dier naar achteren naar de vloer. Veeg de buik met 70% alcohol oplossing. Lokaliseren middellijn van het dier en kwadrant rechtsonder. Spuit de EDE / zoutoplossing in het dier in de onderste rechter kwadrant. Injecteren muizen met Edu / zoutoplossing en het tijdstip van de dag vast te leggen. Herhaal stap 1,2 tot 1,4 op hetzelfde tijdstip van de dag, gedurende 6 opeenvolgende dagen. Opmerking: Als edu negatieve controle nodig is om de gating voor flowcytometrische analyse ingesteld, injecteert tegen leeftijd, geslacht en experimenteel overeenkomened muis met een equivalent volume zoutoplossing zonder Edu. Dit zal de benodigde controle (geen EDU control) te verstrekken. Deze controle moet worden opgenomen voor elk onderzoek om variaties in batches reagentia en cytometer ingesteld. 2. Oogsten Hearts Na 24 uur na het 6e EDE injectie, offeren de muizen met behulp van cervicale dislocatie (of een alternatieve Schedule 1 van Binnenlandse Zaken goedgekeurde methode). Plaats opgeofferd dier in liggende positie en maken de huid incisie van het midden buik om het membraan met chirurgische schaar. Snijd het membraan, en houden het borstbeen vanaf de lichaamsholte, gesneden bilateraal aan het hart bloot te leggen. Til het hart licht en snijd de grote bloedvaten van de uitstroom-darmkanaal naar het hart ontleden uit de borstholte. Plaats onmiddellijk elk hart in een afzonderlijke 15 ml centrifugebuis die 10 ml PBS afgekoeld tot 4 ° C. Transfer harten SEPderlijke 10 cm petrischalen, met een scalpel gesneden elke hart in twee in een sagittale richting en was het hart door voorzichtig te knijpen met een pincet. Vervang vervolgens de PBS om zoveel bloed te verwijderen mogelijk te maken. Herhaal deze stap twee keer. Plaats beide delen van elk hart in hetzelfde individu 1,5 ml microcentrifugebuis. Ga verder met stap 3 of als alternatief op te slaan exemplaren -80 ° C zoals beschreven in de stappen 2.9 en 2.10. Met een 25 G naald, maak een gaatje in de microcentrifugebuis deksel om het deksel niet worden geopend door de uitbreiding van gassen wanneer het hart wordt verwerkt. Label elke microcentrifugebuis en plaats in een container van vloeibare stikstof te bevriezen. 3. Kernen dissociatie en Cardiomyocyte Kernen Labeling Opmerking: Deze kernen dissociatie en cardiomyocyten kernen etikettering wordt aangepast van die van Bergmann et al 11 Deze werkwijze alle monsters geïnjecteerd.met Edu en zoutoplossing ingespoten (geen EDU control) negatieve controle. Zie tabel 1 voor het recept van oplossingen nodig is om deze stappen uit te voeren. Voor elk hart te analyseren, plaats 36 ml van 1% BSA / PBS-oplossing in een ultracentrifuge buis, incubeer gedurende 30 min, gooi de oplossing en laat aan de lucht drogen. Opmerking: Deze buis wordt gebruikt in stap 3,6. Plaats individuele bevroren harten op een 10 mm schotel op ijs en gehakt met behulp van een scalpel waardoor de harten te ontdooien tijdens het hakken proces. Plaats gehakt harten in 15 ml Cell Lysis Buffer in een 50 ml centrifugebuis en gehomogeniseerd met een probe homogenisator gedurende 15 seconden bij 25.000 rpm bij kamertemperatuur. Nogmaals 15 ml Cell Lysis Buffer aan de centrifugebuis overgebracht in een 40 ml Dounce met een grote speling stamper. Voer 10 slagen met de stamper en te filteren door middel van een 100 um dan 40 micrometer cel zeef in een 50 ml centrifugebuis. Centrifugeer gedurende 10 min eent 700 xg bij 4 ° C en verwijder het supernatant. Resuspendeer de nucleaire pellet in 25 ml sucrosegradiënt oplossing en laag het celkernen bevattende oplossing boven 10 ml vers sucrosegradiënt oplossing in de ultracentrifuge gereedgemaakte buis 3,1. Centrifugeer bij 13.000 xg gedurende 1 uur bij 4 ° C met een bal dunne buis rotor. Na het centrifugeren, gooi het supernatant en resuspendeer de kernen pellet in 1300 ui kernen opslag buffer in een 1,5 ml microcentrifugebuis en label PCM1. Verwijder 300 pi van de suspensie naar een nieuwe microcentrifugebuis en toe te voegen aan 700 ul van verse kernen opslag buffer. Label deze buis als ISO controle. Add anti-PCM1 bij een uiteindelijke concentratie van 8 ug / ml aan de microcentrifugebuis gemerkte PCM1 en voeg konijn IgG isotype controle-antilichaam bij een concentratie van 8 ug / ml aan de microcentrifugebuis gemerkte ISO control. Incubeer overnacht bij 4 ° C. Na incubatie, centrifugeer bij 700 xg gedurende 10 min, gooi het supernatans vervangen door 1 ml verse kernen opslagbuffer, hersuspenderen en centrifugeren opnieuw voor 10 min bij 700 x g. Verwijder het supernatant en vervangen door 1 ml verse kernen opslagbuffer. Voeg 1 pl F (ab ') 2 fragment van geit anti-konijn IgG (H + L) antilichaam (FITC of gelijkwaardig groene fluorescerende kleurstof) tot een eindconcentratie van 2 ug / ml te bereiken. Opmerking: Het gebruik van een F (ab ') 2 fragment antilichaam vermindert de kans op niet-specifieke labeling van immuuncellen zoals macrofagen en B-cellen. Wrap buizen aluminiumfolie ter bescherming tegen licht en incubeer gedurende 1 uur bij 4 ° C. 4. Detectie van Edu Incorporation Verdun 10x geconcentreerd Edu reactie buffer 01:10 met gedemineraliseerd water. Centrifuge de PCM-1 en ISO controle gelabelde kernen schorsingen bij 700 g gedurende 10 min, gooi het supernatant en resuspendeer de kernen pellet in 1 ml1% BSA / PBS oplossing. Was de kernen pellet tweemaal. Na de laatste wasbeurt, gooi het supernatant en te vervangen door 100 ul van EDU fixatief. Wikkel buizen aluminiumfolie ter bescherming tegen licht en incubeer bij kamertemperatuur gedurende 15 min. Was de monsters tweemaal met 1 ml van 1% BSA / PBS oplossing, centrifugeer bij 700 xg en incubeer met 1x saponine gebaseerde permeabilisatie oplossing bij kamertemperatuur gedurende 15 min. Bereid 1x Edu etikettering cocktail (tabel 2). Minimaal 2 reacties nodig zijn om de controle geen EDU omvatten. Voeg 500 ul van 1 x EDU etikettering cocktail direct aan elk monster reeds kiembevattende in 100 pl 1x saponine gebaseerde permeabilisatie oplossing en incubeer bij kamertemperatuur beschermd tegen licht gedurende 30 minuten. Centrifugeer bij 700 xg en resuspendeer in 1 ml van 1% BSA / PBS en herhaal deze stap nog twee keer. Centrifuge bij 700 xg, verwijder supernatant en te vervangen door 400 ul van DNA-kleuring-oplossing (zie tabel of Materials). Naam van het reagens 1. Cell Lysis Buffer 0,32 M sucrose 10 mM Tris-HCl (pH = 8) 5 mM CaCl2 5 mM magnesiumacetaat 2,0 mM EDTA 0,5 mM EGTA 1 mM DTT 2.Sucrose Gradient Solution 2,1 M sucrose 10 mM Tris-HCl (pH = 8) 5 mM magnesiumacetaat 1 mM DTT 3. Kernen opslagbuffer (NSB) 0,44 M sucrose 10 mM Tris-HCl (pH = 7,2) 70 mM KCl 10 mM MgCl2 1,5 mM spermine Tabel 1: Kernen isolatie buffer ingrediënten Recept voor alle Buffers worden gebruikt in protocol deel 3 (Kernen dissociatie en cardiomyocyten kernen etikettering).. 5. Flow cytometrische analyse Opmerking: Voer Flowcytometrie op geïsoleerde kernen zoals eerder 7,11,15 beschreven. Maak eerst een perceel beschrijft Forward Scatter (FSC) en zijwaartse verstrooiing (SSC) aan de discriminatie van de kernen van puin (Figuur 2A) mogelijk te maken. Maak een complot om enkele kernen van aggregaten (figuur 2B) discrimineren door het uitzetten 4 ', 6-diamidino-2-phenylindole) (DAPI) gebied (3-405 / 450/50-A) versus de hoogte (33-405 / 450 / 50-H). Zorg ervoor that de 3-405/450/50-A-signaal wordt verzameld op de lineaire schaal naar de juiste bepaling van DNA-inhoud toe te staan. Maak een plot beschrijft 488/535/30 A (PCM-1) vs SSC en slechts gebeurtenissen uit de singlet poort gecreëerd in 5.2 weer te geven met het oog op PCM-1 expressie te evalueren in de singlet bevolking. Voer het isotype controle monster met het oog op een PCM-1 positieve gate (figuur 2C) vast te stellen. Voer een kleine hoeveelheid PCM1 gelabeld monster tot PCM1 verifiëren expressie bevolking en gating positie. Maak een perceel beschrijven SSC-A versus 405/450/50-A (tot DAPI te detecteren). In deze plot, alleen weergave singlet PCM1 uitdrukken kernen, met behulp van de poorten die in 5.3. Gebruik dit complot om een extra hiërarchische poort die alleen 2N kernen (figuur 2D) bevat. Maak een plot dat beschrijft 488/535/30 A vs 1-633/660/20-A tot de identificatie van Edu etikettering van het PCM1 uitdrukken cardiomyocyt bevolking mogelijk te maken. Enkel weergeven kernen die singlet, P zijn CM1 + 2N met de bovenstaande gating. Ren hart monster van geen EDU controle om de EDU positieve gate (figuur 2E) in te stellen. Maak de juiste gate voor EDU + cellen. Record en kwantificeren singlet, PCM-1 uitdrukken, 2N kernen die Edu hebben opgenomen. Bereken deze populatie als percentage van de totale singlet, PCM1 + kernen. Dit zal de mate van neo-cardiomyocyt nucleatie verschaffen gedurende de 7 dagen EDU pulsperiode als percentage van de totale cardiomyocyt kernen. Opmerking: Als DAPI binding aan DNA stoichiometrische de fluorescentie-intensiteit evenredig is met de hoeveelheid DNA. Ploïdie bepalen op basis van de intensiteit van de 405/450/50 A-signaal, zoals hierboven beschreven 7. Om de ploïdie binnen het totale cardiomyocyt bevolking te evalueren gebruik maken van de plot in 5.5 opgericht en een gating strategie gebaseerd op DNA-concentratie 7. /files/ftp_upload/53979/53979fig2.jpg "/> Figuur 2:. Gating strategie om cardiomyocyt Edu oprichting en ploïdie kwantificeren (A) Kernen worden onderscheiden van puin op basis van voorwaartse verstrooiing (FSC) en zijwaartse verstrooiing (SSC). (B) een lineaire poort gemaakt en de singlet kernen populatie is geïdentificeerd op basis van DAPI etikettering en 405/450/50 hoogte vs area signaal. (C) Fluorescerende gating maakt de scheiding van cardiomyocyt kernen (PCM-1-positief) en niet-cardiomyocyt (PCM-1-negatieve) kernen van hartweefsel. (D) Intensiteit van DAPI signaal wordt gebruikt om DNA-concentratie en aldus kernen ploïdie van de PCM1 + cardiomyocyten populatie te bepalen. In muizen> 80% cardiomyocyt kernen diploïde (2n). (E) Fluorescent venstertijd maakt de scheiding van 2N, cardiomyocyt kernen die Edu (PCM + / EDU + = 0,9%) hebben opgenomen van 2N, hartspiercellen die niet hebben opgenomen Edu (PCM + onderwijs). Alle stappen worden beschreven in het protocol 5.0. Zo blijkt uit MDX -. / Y muizen op de C57 / BL10 achtergrond Klik hier om een grotere versie van deze figuur te bekijken.