Direct generation of human induced neurons from human pluripotent stem cells
It is very important that the starting human pluripotent stem cells exhibit a high degree of pluripotency for successful generation of iNs or iOPCs/iOLs. Therefore, cells should be stained for specific markers, such as Oct4 and SOX2, before starting either of the induction protocols described in the present manuscript (Figure 1A). Human H1 cells were used to obtain induced excitatory forebrain neurons following the previously published protocol by Zhang et al. with some modifications (Figure 1C)12,16,17,18. Here, we present a protocol in which iNs at Day 2 are re-plated in pure culture on matrix solution (see Table of Materials), in the absence of any feeder layer: glia or fibroblasts. In addition to the previously published protocols, we observe that freezing iNs at Day 2 does not significantly affect cell viability (~15%–20% cell death after thawing). Pure neurons in culture will start expressing synapsin1 at Day 14–16 (Figure 1D). Establishing a pure neuronal culture is very important because certain factors, for example, the leading AD risk factor ApoE, can be expressed by cells in the feeder layer and this can significantly confound the results.
iOPC generation and iOL maturation is improved by DMSO treatment
Here, we present a fast and efficient protocol that enables the generation of iOPCs in 2 weeks and mature iOLs in 4–5 weeks (Figure 2A). We leveraged the method of transient DMSO treatment we previously developed to augment the differentiation efficiency for ES and iPS cells29,30,31. DMSO treatment enriches the number of cells in the early G1 phase for better signaling integration, favoring differentiation. We performed the first treatment before inducing human ES cells to generate NPCs, and the second treatment before differentiating NPCs into iOPCs. We can detect specific OPC markers (Olig2, CSPG4, NKX2.2, and PDGFRA) as early as 2 weeks after plating of ES H1 cells (Figure 2B,E). The iOPC population at this stage is fairly homogenous, with >95% of cells positive for O4 staining and a high level of immunoreactivity for other markers (Figure 2B). After the start of OL maturation at Day 15, we can typically detect specific OL markers (MBP, O1, CLDN11, and PLP1) starting at Day 28 (Figure 2C,E). The expression of these stage-specific markers correlates with the developmental course of oligodendroglial cells and suggests an accelerated pace, with the NPC markers progressively going down, OPC markers peaking around the second week, and OL markers elevating by the third week (Figure 2D,E)37. Please note that this maturation process diversifies the cell populations. The subpopulations in continuum, comprising multiple intermediate stages between OPCs and mature myelinating oligodendrocytes, can be present and account for varying percentage of total cells, with more mature cells dominating at a later time.
As a comparison, we purchased the highly referenced iOPCs, and matured them in iOL following the manufacturer protocol. We tested the expression of the markers mentioned above in both our iOPC and iOL preparations, and in the cells we purchased. We determined that the cells generated following our protocol had higher expression of all the genes tested (Figure 2E). Interestingly, when we tested the secreted levels of two major isoforms of amyloid-β (Aβ40 and Aβ42) in iNs versus iOPCs, we noticed that iOPCs secreted more of both fragments, but the ratio remained the same (Figure 2F).
Co-culturing of iNs and iOPCs
This protocol is optimized specifically for co-culturing iNs and iOPCs and allow our real-time monitoring of the inter-cellular communications between these two cell types along the course of neural development. The ideal plating densities for both cell types need to be decided with a series of cell number titration to achieve proper differentiation (Figure 3A). After 4 weeks in co-cultures, the iOPCs are expected to be adequately differentiated into OLs that are positive for specific markers such as MBP and extend processes to ensheath axons (Figure 3B). The co-culture system can robustly boost up the number of synapses, indicating that the iOPCs provide a neuronal support through physical contacts or release of trophic factors (Figure 3C). We can maintain the co-cultures in acceptable health condition for up to 6 weeks and observe that the synapse number and other neuronal attributes plateau around the fifth week. Of note, astrocytes and microglia are not present in our preparations and their absence can be documented by checking the expression of specific markers (Figure 3D). The iOPCs express a good number of well-characterized genes that can potentially respond to and mediate the activity-dependent signals from neighboring neurons, in a paracrine (e.g. neurotrophins and metabolites) and/or a synaptic manner (Figure 3E and 3F).
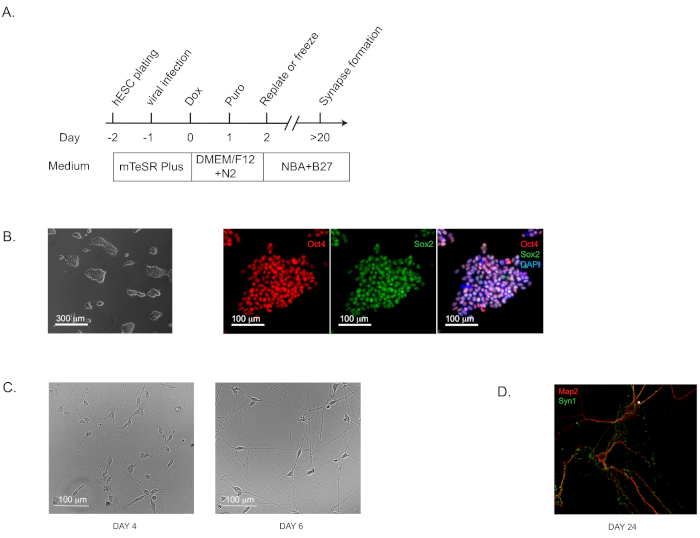
Figure 1: Direct Generation of human induced neurons (iNs) from hPSCs. (A) Flow diagram of iN generation. (B) Representative bright field and immunofluorescence images of the starting culture of human pluripotent stem cells (H1) to confirm the pluripotency. Oct4 is shown in red and Sox2 in green. (C) Representative bright field images of iNs at Day 4 and Day 6. (D) The characteristic morphology for dendritic arborization and synapse puncta in iNs grown in pure culture for 24 days and stained by immunofluorescence staining for dendritic marker Map2 and pre-synaptic marker Synapsin 1 (Syn1). Please click here to view a larger version of this figure.
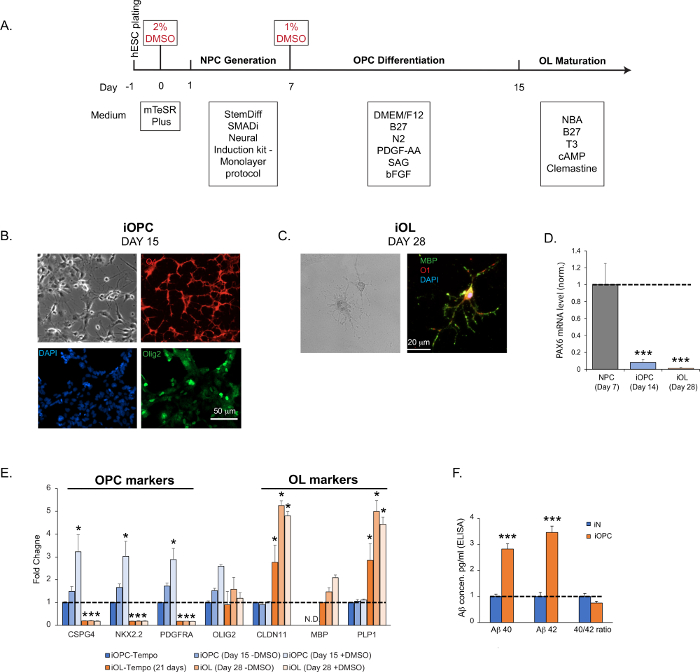
Figure 2: iOPC generation and iOL maturation. (A) Flow diagram of iOPC and iOL generation. (B) Representative bright field and immunofluorescence images of iOPCs at Day 15. Olig2 (pan-oligodendroglia marker) is shown in green, O4 (OPC marker) in red, and DAPI in blue. The imaging revealed that >95% of iOPCs are positive for O4 and 25% for Olig2. (C) Representative bright field and immunofluorescence images of iOLs at Day 28. MBP is shown in green, O1 in red, and DAPI in blue. (D) The expression of NPC marker PAX6 diminishes dramatically in iOPCs at Day 14 and further lowers to background in OLs at Day 28, indicating a robust NPC trans-differentiation and a high level of homogeneity in the iOPC population. (E) The time-course expression profile of common OPC and OL marker genes in cultures generated by the described protocol, without (-DMSO) or with (+DMSO) the step of DMSO incubation (steps 2.1.3 and 2.2.2), assayed at different time points. As a comparison, commercial iOPCs (see Table of Materials) were matured according to the manufacturer’s instructions, and both iOPCs (iOPC-Tempo) or iOLs (iOL-Tempo) were tested for the same markers. As expected, MBP (a mature oligodendrocyte marker) was not detected (N.D.) at the early stages of differentiation in all the iOPCs tested. The DMSO significantly enhanced the efficiency of OPC differentiation and OL maturation. (F) The production and secretion of Aβ40 and Aβ42 in pure iNs and iOPCs cultures, measured by commercial ELISA kits (see Table of Materials) on supernatant obtained from pure iNs and iOPCs cultures both at Day 15 and normalized by cell numbers (both at the density of 200,000 cells per well in a 24-well plate). Please click here to view a larger version of this figure.
Data in bar graphs are plotted as mean ± SEM (n ≥ 3). Statistical significance was evaluated by Student’s t-test (*, p < 0.05; ***, p < 0.001); in (D), compared to the NPC; in (E), compared to the control iOPC-Tempo; in (F), compared to iN.
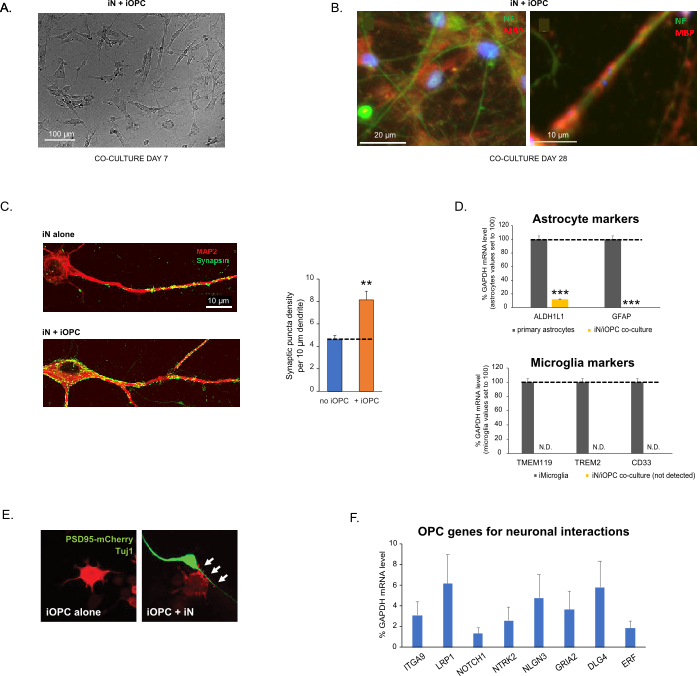
Figure 3: Co-culture of iNs and iOPCs. (A) Representative bright field image of co-cultured iNs and iOPCs at Day 7, showing a proper density for further maturation. (B) Representative immunofluorescence image of iNs and iOPCs co-cultured for 28 days. Axonal marker neurofilament NF is shown in green and oligodendrocytic marker MBP in red. Right, a segment of iN axon ensheathed by iOL process (MBP+). (C) Synapse formation assayed in 4-week-old co-cultures. Cells were stained for Synapsin 1 (Syn1, green) and MAP2 (red), and synaptic puncta were quantified by confocal analysis of density along the dendritic segments as described17,18. (D) In our co-cultures of iNs and iOPCs (7 days of co-culturing), the expression of astrocyte markers, ALDHL1 and GFAP, is minimal (top), and the expression of microglia markers, TMEM119, TREM2, and CD33, is not detected (N.D.) by qPCR. The contamination from these two glial cell types is thus excluded. (E) Coculturing iOPC with iN leads to the formation of neuron-OPC synapses. The fluorescence-tagged post-synaptic marker PSD95-mCherry is expressed only in OPCs, and display a diffuse pattern in single cultures (left) but aggregate to form puncta in cocultures (right, indicated by arrows; Tuj1, neuronal marker). (F) The expression of well-characterized oligodendroglial genes that can sense and respond to neuronal activities in the pure cultures of iOPCs at Day 14. Please click here to view a larger version of this figure.
Data in bar graphs is plotted as mean ± SEM (n ≥ 3). Statistical significance was evaluated by Student’s t-test (**, p < 0.005; ***, p < 0.001); in (C), compared to the no OPC condition; in (D), compared to primary astrocytes in top panel.
