La determinación de la estructura de una biomolécula es un requisito clave para la comprensión de su función. Dos métodos bien establecidos para la determinación de la estructura son crio-microscopía electrónica y cristalografía de rayos X 1, 2. Hoy en día, ambos métodos proporcionan información estructural de alta resolución con una resolución hasta el nivel de angstrom. Estos dos métodos se han utilizado ampliamente para elucidar la estructura de las grandes biomoléculas tales como los complejos de proteínas. Aunque los métodos existentes constantemente se han mejorado a lo largo de las últimas décadas, la complejidad de las estructuras biológicas sigue planteando un desafío importante para la biología estructural, en particular cuando grandes complejos, dinámicos y transitorios son investigados 3.
Con el fin de estudiar la dinámica de complejos macromoleculares y la relación estructura-función en particular, las metodologías de una sola molécula tienen provided información útil 4. Se desarrollaron varias nuevas estrategias para afrontar de manera ortogonal en la adquisición de información estructural y dinámica. Ejemplos son altos AFM velocidad 5, 6 manipulación mecánica, fluorescencia localización microscopía 7, así como de una sola molécula de transferencia de energía de resonancia de Förster (smFRET) 8, 9. Desde muy temprano en FRET que se ha denominado una regla molecular, debido a la dependencia de la distancia en la escala de longitud de biomacromoléculas 10.
Una aplicación particularmente interesante de smFRET es utilizar la información de distancia obtenida a partir de mediciones smFRET para inferir información estructural 11, 12, 13, 14, 15 </sup>, 16, 17, 18, 19, 20, 21, 22, 23. Debido a la alta resolución temporal de smFRET, la posición de las partes móviles de una estructura de proteína puede ser localizada. Sin embargo, con el fin de extraer información cuantitativa a partir de datos smFRET importantes parámetros de corrección sobre las moléculas de colorante que determinarse durante la medición 24. Con estos factores de corrección, la eficiencia de FRET FRET E se puede calcular utilizando la fórmula
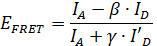 ,
,
en la que A y D I </sub> son las intensidades de fluorescencia del donador y la molécula aceptora, respectivamente (ver Figura 2). El β-factor de cuenta de la diafonía, la fuga de emisión del donante en el canal aceptor y se calcula

donde I 'A e I' d son las intensidades de fluorescencia del donador y la molécula aceptora después de la foto de blanqueo de la molécula aceptora.
El γ-factor de corrige la diferencia en las eficiencias de detección relativos en los dos canales, así como las diferencias en el rendimiento cuántico de fluorescencia del donante y el colorante aceptor. Se calcula a partir de cada curva del tiempo individual
<p class="jove_content" fo:keep-together.within-page="1" fo:text-align="center" style = "text-align: center;">
Nota, que esta descripción descuida excitación directa de la molécula aceptora, que a veces se vuelve importante y tendría que ser corregida para así. Para la determinación de estos factores de corrección es útil para excitar tanto para el donante, así como el aceptor en un esquema de alternancia 25 con el fin de diferenciar entre los cambios de foto-física y dinámica estructural.
Con el fin de no sólo lograr eficiencias smFRET cuantitativos, sino también información estructural cuantitativa, se introdujo el Sistema de Posicionamiento Nano-(NPS) en 2008 26. El nombre fue elegido en base a sus similitudes con el sistema de posicionamiento global por satélite (GPS). El NPS es una técnica híbrida que combina smFRET y datos de cristalografía de rayos X para la localización de las posiciones de tinte desconocidos en los complejos biomacromoleculares. La Cestructura rystal sirve como un marco de referencia y los resultados smFRET se utilizan para obtener información de la distancia entre una posición desconocida fluoróforo (antena) y una posición conocida de la estructura cristalina (satélite). En experimentos consecutivos de las distancias entre la antena y varios satélites se miden y la posición de la antena se determina por medio de un esquema de análisis estadísticamente rigurosa basada en la estimación de parámetros Bayesiano. Como resultado, no sólo la posición más probable de la antena se calcula, pero su distribución de la incertidumbre en 3D completa, la llamada posterior, visualizado por volúmenes creíbles. Por otra parte, fuentes de energía nuclear fue ampliado para permitir el análisis de redes completas smFRET 27.
El NPS se ha utilizado para resolver una serie de cuestiones importantes de la transcripción eucariótica, es decir, el curso de la DNA aguas arriba, el ADN no molde y el ARNm naciente dentro de la elongación co RNA polimerasa IImplex 12, 28, demuestra también el efecto de los factores de transcripción iniciación 26 y la arquitectura dinámica de un promotor-abierta compleja 29. Además, el NPS se utilizó para elucidar la estructura de la archaeal complejo abierto ARN polimerasa 30 y, en particular, la posición del factor de iniciación de la transcripción TFE, que se une competitivamente al mismo sitio como el factor de elongación de la transcripción SPT4 / 5 31.
Desde entonces, una serie de enfoques estructurales basados smFRET se han publicado 15, 18, 21, 23. Al comparar los diferentes métodos estructurales basados smFRET, se hace evidente que la aparente precisión del método depende de la elección particular de los modelos de colorantes altamente. Hay que señalar quemoléculas de colorante pueden mostrar un comportamiento espacial y de orientación diferente en función de su entorno local.
Con este fin, Fast-NPS se introdujo 32. Fast-NPS usa un algoritmo de muestreo avanzado reducir los tiempos de cálculo drásticamente. Por otra parte, Fast-NPS permite a uno realizar un análisis estructural y para cada molécula de colorante que el usuario puede elegir entre una serie de cinco modelos distintos de colorantes que se describirá a continuación. El modelo más conservador, denominado clásico, asume que el colorante ocupa sólo una, pero desconocida, posición. En esta posición, el fluoróforo puede girar libremente dentro de un cono, cuyo tamaño está determinado a partir de su respectivo anisotropía de fluorescencia (dependiente del tiempo). La orientación del cono no es conocida, lo que conduce a grandes incertidumbres al convertir eficiencias smFRET medidos en distancias. A este respecto, el modelo es conservadora, ya que conducirá a la precisión más pequeño en comparación con el otro modo de colorantels. Sólo para distancias muy cortas caso de que las suposiciones hechas por el modelo clásico de plomo a una determinación de la posición notoriamente incorrecto. Para valores típicos smFRET, la posición correcta se entrega siempre en el volumen comparativamente grande creíble.
Sin embargo, desde una precisión más alta es deseable, es importante desarrollar y probar modelos alternativos de colorantes, que podrían ayudar a mejorar la precisión. Si el colorante gira mucho más rápido que su tiempo de vida de fluorescencia inherente, el llamado modelo ISO se puede aplicar. Aquí, el factor de orientación kappa 2 (necesaria para el cálculo del radio de Förster característica isotrópica  ) Está ajustado a 2/3. Como resultado, los volúmenes creíbles calculadas son casi dos órdenes de magnitud más pequeña en comparación con aquellos en el modelo clásico de 32. En el caso de que el fluoróforo se encuentra en un entorno que permita no sólo reori rápidoentación, pero, además, el movimiento rápidamente en todo su volumen accesible, el modelo meanpos-iso debe ser utilizado. En este modelo, el tinte ocupa efectivamente sólo una posición media, en el que el promedio espacial se explica por una conversión distancia polinomio 15. Este modelo se aplica si, por ejemplo el colorante (comúnmente hidrófobo) se une a una región hidrófila, por ejemplo, el ADN. La aplicación del modelo meanpos-iso conduce a una reducción adicional en el tamaño de los volúmenes creíbles por un factor de aproximadamente dos. Sin embargo, un colorante unido a una proteína puede unirse de forma reversible a varios parches hidrófobos en su volumen estéricamente accesible (AV). Un fluoróforo que conmuta instantáneamente entre estas regiones, pero dentro de una región sufre una rotación libre y el movimiento localizada rápido es mejor descrita por el modelo VAR-meanpos-iso. Para una situación similar en la que el colorante no es libre de girar las var-meanpos aplica modelo. más d etails acerca de estos modelos se pueden encontrar en nuestra reciente publicación 32.
) Está ajustado a 2/3. Como resultado, los volúmenes creíbles calculadas son casi dos órdenes de magnitud más pequeña en comparación con aquellos en el modelo clásico de 32. En el caso de que el fluoróforo se encuentra en un entorno que permita no sólo reori rápidoentación, pero, además, el movimiento rápidamente en todo su volumen accesible, el modelo meanpos-iso debe ser utilizado. En este modelo, el tinte ocupa efectivamente sólo una posición media, en el que el promedio espacial se explica por una conversión distancia polinomio 15. Este modelo se aplica si, por ejemplo el colorante (comúnmente hidrófobo) se une a una región hidrófila, por ejemplo, el ADN. La aplicación del modelo meanpos-iso conduce a una reducción adicional en el tamaño de los volúmenes creíbles por un factor de aproximadamente dos. Sin embargo, un colorante unido a una proteína puede unirse de forma reversible a varios parches hidrófobos en su volumen estéricamente accesible (AV). Un fluoróforo que conmuta instantáneamente entre estas regiones, pero dentro de una región sufre una rotación libre y el movimiento localizada rápido es mejor descrita por el modelo VAR-meanpos-iso. Para una situación similar en la que el colorante no es libre de girar las var-meanpos aplica modelo. más d etails acerca de estos modelos se pueden encontrar en nuestra reciente publicación 32.
Estos modelos ofrecen un amplio repertorio para dar cuenta específicamente para los diferentes ambientes un tinte podría encontrar y aplicar de manera inteligente optimiza la precisión de localización. En Fast-NPS cada molécula de colorante unido a una posición específica se puede asignar a un modelo individual, de tal manera que FRET-socios se les permite tener diferentes modelos. Esto permite el modelado ilimitadas y cerca de la naturaleza. Sin embargo, es importante que uno realiza pruebas estadísticas rigurosas para asegurar que el resultado obtenido por la combinación final modelo es todavía de acuerdo con los datos experimentales. Estas pruebas están incluidas en el software Fast-NPS.
Para la aplicación de Fast-NPS a los datos experimentales se requiere la medición de (sólo) tres parámetros de entrada. En primer lugar, el tinte-par específico Förster isotrópica radios (/54782/54782eq5.jpg "/>) Se han determinado. Por lo tanto, el rendimiento cuántico (QY) del colorante donante, los espectros de emisión de fluorescencia del donante y el espectro de absorción del aceptor deben ser medidos. Estas mediciones pueden llevarse a cabo en granel, usando un espectrómetro estándar y un espectrómetro de fluorescencia estándar. Para cada par, la R 0 se calcula entonces utilizando la PhotochemCAD freeware y se puede utilizar en el análisis de NPS. Por otra parte, los (resuelta en el tiempo) anisotropías de fluorescencia de las moléculas de colorante necesitan ser obtenido usando una polarización (y tiempo) espectrómetro de fluorescencia sensible. Sin embargo, los parámetros de entrada más importantes para Fast-NPS son las eficiencias smFRET medidos en una configuración de microscopía de fluorescencia de una sola molécula, tales como un microscopio de fluorescencia de reflexión total interna (TIRFM) .
A continuación, presentamos un protocolo paso a paso para la obtención de datos y la aplicación de smFRET Fast-NPS (Figura 1).
