Reaktif oksijen türleri (ROS) sinyali sinir sisteminin gelişimini ve işleyişini düzenler1. Önemli bir hücresel ROS kaynağı, süperoksit ve hidrojen peroksit (H2O 2 )2üreten transmembran proteinleri olan NADPH oksidazlarıdır (NOX). NOX enzimleri merkezi sinir sistemi (CNS) boyunca bulunur ve NOX türevi ROS nöronal gelişime katkıda bulunur3,4,5,6. Sinirsel kök hücrelerin bakımı ve farklılaşması, nöronal polaritenin kurulması, neurit büyümesi ve sinaptik plastisitenin yeterli ROS7, 8,9,10,11seviyelerini gerektirdiği gösterilmiştir. Öte yandan, NOX’ler tarafından kontrolsüz ROS üretimi Alzheimer Hastalığı, multipl skleroz ve travmatik beyin hasarı dahil olmak üzere nörodejeneratif bozukluklara katkıda bulunur12,13,14. Bu nedenle, fizyolojik olarak ilgili ROS üretimi sağlıklı koşulların korunması için kritik öneme sahiptir.
Genetik olarak kodlanmış biyosensörlerin geliştirilmesi hücresel ROS’un tespitini büyük ölçüde kolaylaştırmıştır. Genetik olarak kodlanmış biyosensörlerin önemli bir avantajı, ROS sinyalinin artan zamansal ve mekansal çözünürlüğüdür, çünkü bu sensörler özellikle farklı konumlara hedeflenebilir. Redoks duyarlı GFP (roGFP) bu tür ROS biyosensörlerinin bir türüdür. RoGFP2-Orp1 varyantı özellikle Maya15,16’dan glutatyon peroksitoksin aile proteini olan Orp1 etki alanı aracılığıyla H 2 O2’yi algılar. Orp1 proteininin oksidasyonu, konformasyonunu değiştirmek için roGFP2’ye aktarılır (Şekil 1A). Prob, 405 nm ve 480 nm’ye yakın iki heyecan zirvesi ve 515 nm’de tek bir emisyon zirvesi sergiliyor. Oksidasyon üzerine, heyecan zirvesi etrafındaki floresan yoğunluğu değişir: 405 nm heyecan artarken, 480 nm heyecan azalır. Bu nedenle, roGFP2-Orp1 oranmetrik bir biyosensördür ve H2O2-seviyeleri iki farklı dalga boyunda heyecanlanan floresan yoğunluklarının oranı ile tespit edilir (Şekil 1B). Genel olarak, roGFP2-Orp1, ROS görüntüleme için verimli bir şekilde kullanılabilecek çok yönlü bir araçtır vivo.
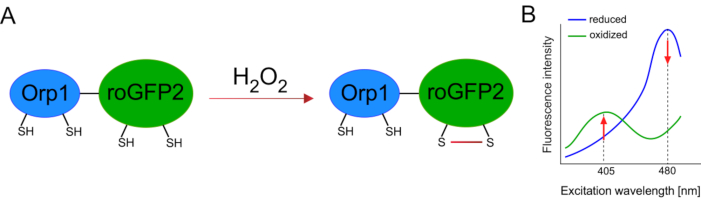
Şekil 1: RoGFP2-Orp1’in şematik gösterimi ve eksitasyon spektrumları. (A) Oksidan transferi, H 2 O 2’ye yanıt olarak Orp1 ve roGFP2 arasındagerçekleşirve roGFP2’de konformasyon değişikliklerine yol meydana gelir. (B) RoGFP2-Orp1’in heyecan spektrumu, 405 nm ve 480 nm’de iki heyecan zirvesi ve 515 nm’de tek emisyon zirvesi sergiler. H 2 O2ileoksidasyonüzerine, 405 nm ekscitasyon artarken 480 nm heyecan azalır. Bu, H2O2 varlığı için oranmetrik bir okuma ile sonuçlanır. Rakam Bilan ve Belousov (2017)16 ve Morgan ve ark. (2011)25‘ten değiştirilmiştir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
Danio rerio (zebra balığı) model sistemi, genetik olarak kodlanmış biyosensörleri uygulamak için çeşitli avantajlara sahiptir. Embriyoların ve larvaların optik şeffaflığı invaziv olmayan in vivo görüntülemeyi mümkün hale sağlar. Daha yüksek çözünürlük ve daha derin penetrasyon elde etmek için yeni görüntüleme araçları geliştirilmektedir17. Ayrıca, transgenik hayvanların neslini destekleyen genetik manipülasyon (ektopik mRNA ekspresyözü, Tol2 transgenez, vb.) ve genom düzenleme (TALENler, CRISPR / Cas9, vb.) için yerleşik araçlar vardır18. Zebra balığı embriyoları anne dışında geliştikçe, bu sistem embriyoların daha kolay erişmesini ve manipülasyonuna izin verir. Örneğin, tek hücreli aşama enjeksiyonları ve ilaç tedavileri kolayca yapılabilir.
Burada zebra balığını, H 2 O2’yeözgü biyosensör roGFP2-Orp1’i tüp bebekli mRNA enjekteederekgeçici olarak ifade etmek için kullandık. Bu embriyolar hem kültürlü nöronların in vitro görüntülemesi hem de in vivo görüntüleme için kullanılabilir ( Şekil2). Zebra balığı embriyolarından retinal ganglion hücrelerinin (RGC) parçalanıp kaplatılmasını ve ardından kültürlü nöronlarda H2O2seviyelerinin değerlendirilmesini içeren bir protokol açıklıyoruz. Daha sonra konfokal mikroskopi kullanarak roGFP2-Orp1 ekspresyaklı embriyo ve larvaların in vivo görüntüleme yöntemi sunuyoruz. Bu yaklaşım sadece fizyolojik H2O2seviyelerinin belirlenmesine izin vermekle kalmaz, aynı zamanda farklı gelişim aşamalarında veya koşullarında meydana gelen potansiyel değişiklikleri de belirler. Genel olarak, bu sistem H 2 O2’ningelişim, sağlık ve hastalıktaki rolünü incelemek için canlı hücrelerde ve hayvanlarda H2O2’yi tespit etmek için güvenilir bir yöntem sağlar.

Şekil 2. Deneysel yaklaşımın ana hatları. Kısaca embriyo toplanmasından sonra roGFP2-Orp1 mRNA tek hücreli evre zebra balığı embriyolarının sarısına enjekte edilir. Gelişmekte olan embriyolar hem (A) in vitro hem de (B) in vivo görüntüleme için kullanılabilir. (A) GFP pozitif embriyolar 34 hpf’de RGC koleksiyonu için retinaları parçalamak için kullanılır. Ayrışmış RFC’ler ZFCM (+) ortamlarında PDL/laminin kaplı coverlips üzerine kaplanmıştır. RGC’ler 6-24 saat kaplamadan sonra aksonlarını uzattıkça büyüme konisi görüntülemesi yapılabilir. Hücreler, H2O2seviyelerindeki olası değişiklikleri ölçmek için farklı tedavilere tabi tutulabilir. Burada RGC’lerin (kırmızı) büyüme konilerindeH2 O2seviyelerini ölçtük. (B) In vivo görüntüleme için GFP pozitif embriyolar kullanılır. İstenilen yaşta embriyolar uyuşturulabilir ve konfokal görüntüleme için 35 mm cam dip yemeklerine monte edilebilir. Burada embriyolar retina görüntüleme için ventral olarak monte edilir. Şematik zebra balıklarında retina gelişimini gösterir. RGC’ler retinadaki en iç katman olan ganglion hücre tabakasını (GCL) oluşturur. RGC aksonları orta çizgiyi geçmek için optik sinire dönüşür ve optik chiasm oluşturur. Daha sonra, RGC aksonları, orta beyindeki optik tectum’da sinaps yapmak için dorsally büyür. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
