חמצן תגובתי מינים (ROS) איתות מווסת את ההתפתחות והתפקוד של מערכת העצבים1. מקור ROS תאי חשוב הם תחמוצות NADPH (NOX), שהם חלבונים טרנסממברן המייצרים סופראוקסיד ומי חמצן (H2O2)2. אנזימי NOX נמצאים בכל מערכת העצבים המרכזית (מערכת העצבים המרכזית), ו- NOX נגזר ROS לתרום להתפתחות עצבית3,4,5,6. תחזוקה ובידול של תאי גזע עצביים, יצירת קוטביות עצבית, נוייריט נימים, פלסטיות סינפטית הוכחו לדרוש רמות נאותות של ROS7,8,9,10,11. מצד שני, ייצור בלתי מבוקר של ROS על ידי NOXes לתרום הפרעות ניווניות כולל מחלת אלצהיימר, טרשת נפוצה, ופגיעה מוחית טראומטית12,13,14. לפיכך, הייצור של ROS רלוונטי מבחינה פיזיולוגית הוא קריטי לשמירה על תנאים בריאים.
פיתוח של biosensors מקודדים גנטית הקל על גילוי של ROS הסלולר מאוד. אחד היתרונות החשובים של biosensors מקודד גנטית הוא הרזולוציה הזמנית והמרחבית מוגברת של אות ROS, כמו חיישנים אלה יכולים להיות ממוקדים במיוחד למקומות שונים. GFP רגיש לאדומים (roGFP) הוא סוג אחד של ביוסנסורים מסוג ROS. גרסת roGFP2-Orp1 מזהה באופן ספציפי H2O2 דרך תחום Orp1 שלה, שהוא חלבון משפחת גלוטתיון peroxiredoxin משמרים15,16. החמצון של חלבון Orp1 מועבר roGFP2 כדי לשנות את הקונפורמציה שלה (איור 1A). הגשוש מציג שתי פסגות עירור ליד 405 ננומטר ו 480 ננומטר, ושיא פליטה יחיד ב 515 ננומטר. עם החמצון, עוצמת הפלואורסצנטיות סביב עירור מגיעה לשיאה: בעוד שהתרגשות של 405 ננומטר עולה, 480 ננומטר של עירור פוחת. לפיכך, roGFP2-Orp1 הוא ביוסנסור יחסי, ורמות H2O2מזוהות על ידי היחס בין עוצמות הפלואורסצנטיות הנרגשות משני אורכי גל שונים (איור 1B). בסך הכל, roGFP2-Orp1 הוא כלי רב-תכליתי להדמיית ROS שניתן להשתמש בו ביעילות ב- vivo.
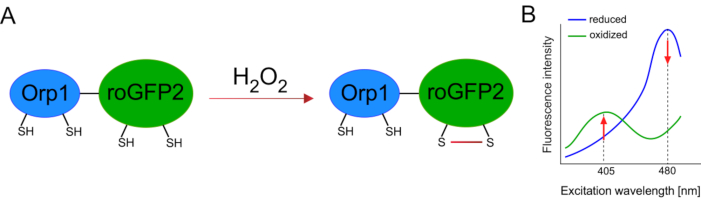
איור 1: ייצוג סכמטי וספקטרום עירור של roGFP2-Orp1. (A) העברת חמצון מתרחשת בין Orp1 ו roGFP2 בתגובה H2O2, המוביל לשינויים קונפורמיים roGFP2. (B)ספקטרום העירור של roGFP2-Orp1 מציג שתי פסגות עירור ב 405 ננומטר ו 480 ננומטר ושיא פליטה יחיד ב 515 ננומטר. עם חמצון על ידי H2O2, עירור 405 ננומטר עולה בעוד 480 ננומטר עירור פוחתת. התוצאה היא קריאה ביחס עבור נוכחות H2O2. הנתון שונה מבילאן ובלושוב (2017)16 ומורגן ואח ‘ (2011)25. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
למערכת המודלים של דניו ריו (זברה-פיש) יש מספר יתרונות ליישום ביוסנסורים מקודדים גנטית. השקיפות האופטית של העוברים והזחלים מאפשרת הדמיה לא פולשנית של vivo. כלי הדמיה חדשים מפותחים כדי להשיג רזולוציה גבוהה יותר וחדירה עמוקה יותר17. יתר על כן, ישנם כלים מבוססים למניפולציה גנטית (ביטוי mRNA חוץ רחמי, Tol2 transgenesis, וכו ‘) ועריכת גנום (TALENs, CRISPR / Cas9, וכו ‘), אשר מקדם את הדור של בעלי חיים מהונדסים18. ככל שעוברי הזברה מתפתחים מחוץ לאם, מערכת זו מאפשרת גישה ומניפולציה קלות יותר של העוברים. לדוגמה, זריקות שלב אחד וטיפולים תרופתיים ניתן לעשות בקלות.
כאן, השתמשנו בזברה כדי לבטא באופן ארעי את ה- H2O2-ספציפי biosensor roGFP2-Orp1 על ידי הזרקת mRNA עם עיטור במבחנה. עוברים אלה יכולים לשמש הן להדמיה במבחנה של נוירונים תרבותיים והן להדמיית vivo (איור 2). אנו מתארים פרוטוקול עבור ניתוח וציפוי תאי גנגליון רשתית (RGCs) מעוברים זברה ולאחר מכן הערכת H2O2-רמות נוירונים תרבותיים. לאחר מכן, אנו מציגים שיטה להדמיה in vivo של עוברים מבטאים roGFP2-Orp1 ותזחלים באמצעות מיקרוסקופיה קונפוקלית. גישה זו לא רק מאפשרת לקבוע פיזיולוגית H2O2-רמות, אלא גם שינויים פוטנציאליים המתרחשים בשלבים התפתחותיים שונים או תנאים. בסך הכל, מערכת זו מספקת שיטה אמינה לגילוי H2O 2 בתאים חיים ובעליחיים כדי ללמוד את התפקיד של H2O2 בפיתוח, בריאות, ומחלות.

איור 2. קווי המתאר של הגישה הניסיונית. בקצרה, לאחר איסוף עוברים, roGFP2-Orp1 mRNA מוזרק לתוך החלמון של עוברי זברה שלב אחד. פיתוח עוברים יכול לשמש הן עבור (A) במבחנה ו –( B) בהדמיה vivo. (A)עוברים חיוביים GFP משמשים לנתח רשתית עבור אוסף RGC ב 34 hpf. RGCs מנותקים מצופים על כריכות מצופות PDL/למין במדיה ZFCM (+). הדמיית חרוט צמיחה יכולה להתבצע כמו RGCs להרחיב את האקסונים שלהם לאחר 6-24 שעות של ציפוי. תאים יכולים להיות כפופים לטיפולים שונים כדי למדוד את השינויים הפוטנציאליים H2O2-רמות. כאן, מדדנו H2O2-רמות בקונוסים הצמיחה של RGCs (אדום). (B)עוברים חיוביים של GFP משמשים להדמיית vivo. בגיל הרצוי, עוברים יכולים להיות מורדמים ומורכבים על כלים תחתונים מזכוכית 35 מ”מ להדמיה קונפוקלית. כאן, עוברים מותקנים באופן מאוורר להדמיה ברשתית. סכמטי מראה התפתחות רשתית בזברפיש. RGCs יוצרים שכבת תא גנגליון (GCL), שהיא השכבה הפנימית ביותר ברשתית. אקסונים RGC לפתח עצב הראייה לחצות קו האמצע, ויוצרים chiasm אופטי. לאחר מכן, אקסונים RGC לגדול dorsally לעשות סינפסות ב טקטום אופטי ב midbrain. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
