BMEC Differentiation
A few critical steps in this protocol should be followed precisely (Figure 1). E6 medium use on day 1 is important, since it is often used for deriving neuroectoderm lineage from iPSCs within a relatively short period of time yielding reproducible results across multiple cell lines36. Another important step is on day 4 of differentiation, where E6 medium should be switched to hESFM with diluted (1:200) B27, 20 ng/mL bFGF and 10 μM RA to expand iPSC-derived BMECs. The addition of B27 supplement is used as an alternative to bovine serum to support serum-free cell culturing1, bFGF is added to facilitate growth of iPSC-derived BMECs6, and RA is used to facilitate the development of the BBB phenotype35. The last important step involves the purification stage, where day 6 iPSC-derived BMECs are sub-cultured onto a COL4/FN coated plate to select iPSC-derived BMECs1,6,35,36. Figure 2 demonstrates the morphological transition from iPSCs to BMECs. After one day of E6 medium (day 1), cellular morphology is similar to that of iPSCs. By day 4 of E6, cells begin to appear visibly distinct from iPSCs and cover most of the well (~90% confluency). By day 6, while cultured in hESFM with diluted (1:200) B27, 20 ng/mL bFGF and 10 μM RA, cellular morphology begins to have an elongated and cobblestone appearance. At day 8, each individual cell is distinct in a large cobblestone pattern. A sprouting assay was performed to demonstrate the angiogenic potential of iPSC-derived BMECs, which resulted in tube-like structures after 3 days of VEGFA165 treatment (Figure 3).
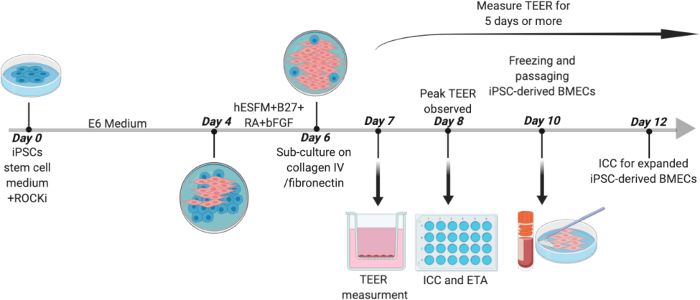
Figure 1: Outline for Differentiation of Human iPSCs to BMECs. Human iPSCs were initially cultured in stem cell medium containing 10 μM Y-27632 for 24 hours before changing medium to E6 for 4 days. On day 4, medium was changed to hESFM with (1:200) B27 supplement, 20 ng/mL bFGF, and 10 μM RA for 2 days. On day 6, cells were sub-cultured onto COL4/FN coated plates. On day 7, medium was changed to hESFM with B27 supplement without bFGF and RA and TEER was measured. On day 8, ICC and efflux transporter activity assays were performed. iPSC-derived BMECs were expanded until day 10 before being passaged to a trans well plate or a 24-well flat bottom plate for TEER measurement and ICC analysis, respectively. Day 8 BMECs were used for the sprouting assay (not depicted). 2 wells of a 6-well plate of iPSC-derived BMECs were collected and stored in hESFM with 10% DMSO and 30% FBS at -80 °C and then in liquid nitrogen for long-term storage at -160oC. On day 12, a peak in TEER value was observed in expanded iPSC-derived BMECs at which point ICC was performed. Please click here to view a larger version of this figure.

Figure 2: Bright-field Images Depicting Differentiation of iPSCs to BMECs. After one day of culture in E6 medium, iPSCs retain their characteristic morphology. On day 4 in E6 medium, cellular morphology appears distinctly different from iPSCs. On day 6, cellular morphology changes to an elongated and cobblestone appearance. By day 8, cells appear large and with a cobblestone pattern. Please click here to view a larger version of this figure.
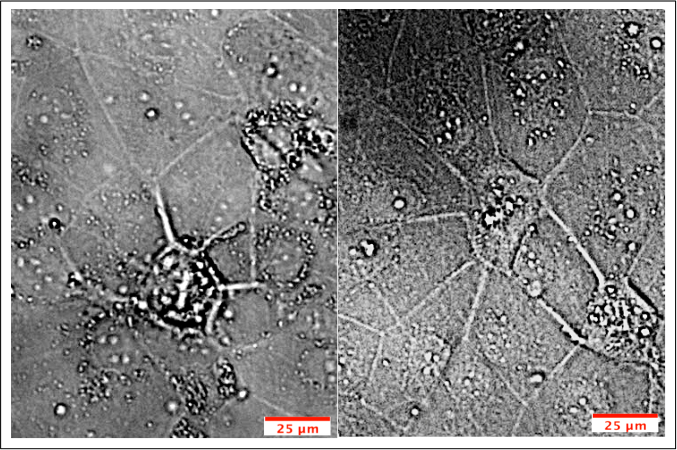
Figure 3: Angiogenic Potential of iPSC-derived BMECs. Purified iPSC-derived BMECs were seeded at 100,000 cell/cm2 onto basement membrane matrix in hESFM with (1:200) B27 supplement and 40ng/mL VEGFA165. Tube-like structures appeared after 3 days of VEGFA165 treatment. Please click here to view a larger version of this figure.
Purified iPSC-derived BMECs were seeded at 100,000 cell/cm2 onto basement membrane matrix in hESFM with (1:200) B27 supplement and 40ng/mL VEGFA165. Tube-like structures appeared after 3 days of VEGFA165 treatment.
BMEC characterization was performed using immunocytochemistry for cell-specific markers. iPSC-derived BMECs were assessed for the presence of tight junction proteins (OCLN, TJP1, and CLDN5), which are commonly expressed in the tight junctions of brain endothelial cells3 and endothelial cells in the lung, liver, and kidney18. Other markers such as PECAM1 and SLC2A1, have been previously used as markers for purified BMECs6. PECAM13 and SLC2A4 are both expressed in vascular endothelial cells of the BBB. The iPSC-derived BMECs generated using this protocol co-expressed all five of these markers (Figure 4).

Figure 4: Marker Analysis of iPSC-Derived BMECs. Human iPSC-derived BMECs were stained for tight junction (OCLN, TJP1, CLDN5), influx transporter (SLC2A1), and adherens junction (PECAM1) proteins. OCLN, TJP1, and CLDN5 proteins are primarily localized in the cell membrane. SLC2A1 and PECAM1 are localized in both the nuclei and cell membrane. Hoechst 33342 trihydrochloride trihydrate was used for nuclear staining. Please click here to view a larger version of this figure.
To characterize BBB function of BMECs, TEER was measured 24 hours (day 7) after sub-culturing and the medium was changed to hESFM with diluted (1:200) B27 without bFGF and RA. TEER measurements were obtained starting at day 7 of differentiation (day 0 of TEER measurement) and peaked at ~2000 Ω x cm2 on day 8 or 48 hours after sub-culturing BMECs (Figure 5). These TEER values are within the range described for co-cultured iPSC-derived BMECs with rat primary astrocytes37. The iPSC line did not have any discernable BBB function according to their low TEER values.
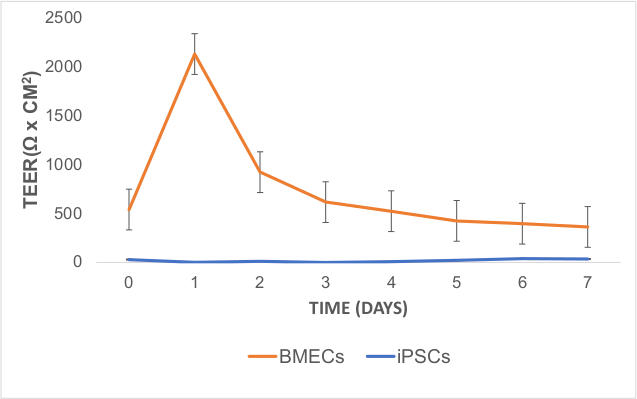
Figure 5: TEER Measurements in iPSC-Derived BMECs. TEER values peaked after one day of sub-culturing on COL4/FN matrix (on day 8 of differentiation). TEER measurements were obtained in technical (3 measures per well) and biological replicates (3 wells per cell line). The technical average value from a blank well was subtracted from raw TEER values. These values were averaged for each day and multiplied by 1.12 cm2 (surface area of the 12-transwell insert). Error bars represent standard error. Please click here to view a larger version of this figure.
To evaluate ABCB1 and ABCC1 efflux transporter activity, the amount of fluorescent substrate taken up for ABCB1 and ABCC1 were quantified following incubation with their respective inhibitors. As expected, inhibition of ABCB1 and ABCC1 efflux transporters with PSC833 (ABCB1 inhibitor) or MK-571 (ABCC1 inhibitor) led to an increase in rhodamine 123 (R123) or 2’,7’-dichlorodihydrofluorescein diacetate (H2DCFDA), respectively (Figure 6). This evidence suggests that BMECs derived using this protocol have efflux transporter activity.
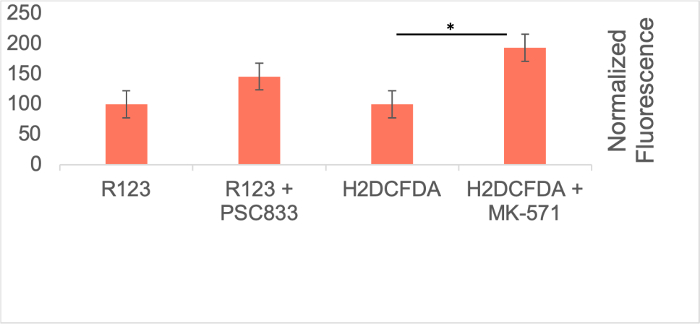
Figure 6: Efflux Transporter Activity in iPSC-Derived BMECs. Efflux transporter activity in BMECs was determined by quantifying the accumulation of rhodamine 123 (R123) or 2',7'-dichlorodihydrofluorescein diacetate (H2DCFDA) in the presence or absence of PSC833 (ATP binding cassette subfamily B member 1 (ABCB1) inhibitor) or MK-571 (ATP binding cassette subfamily C member 1 (ABCC1) inhibitor). Technical triplicates were performed for each condition (N=1). Fluorescence values from the control condition (i.e. without inhibitors) were deducted from raw fluorescence values. These fluorescence accumulation was normalized on a per-cell basis for each technical replicate. Statistical significance was determined using student t-test from the three technical replicates. No statistical significance was observed between the accumulation of R123 with and without ABCB1 inhibitor (t-stat= -1.66, p=0.11). Statistical significance was observed between the accumulation of H2DCFDA with and without ABCC1 inhibitor (t-stat=-7.23, p=0.04). *p<0.05. Error bars represent standard error. Please click here to view a larger version of this figure.
Passaging, Expanding and Cryopreserving iPSC-Derived BMECs
Another aim was to investigate whether iPSC-derived BMECs could be passaged and cryopreserved after sub-culturing. For this purpose, day 7 iPSC-derived BMECs were allowed to expand until day 10 before passaging them onto newly coated COL4/FN 12-transwell filtered plates for TEER measurement and 24-well flat-bottom plate for ICC analysis (Figure 7). Using this condition iPSC-derived BMECs continued to proliferate, maintained the expression of OCLN, TJP1, CLDN5, SCL2A1, and PECAM1 (Figure 8), and continued to sustain proper TEER values (peak at~2000 Ω x cm2) after passaging (Figure 9). Cryopreserved BMECs were later thawed, expanded, and then passaged (Figure 7). TEER measurements of BMECs were obtained 24 hours after thawing and several more days after that. TEER measurements of these post-thawed BMECs were reduced (peaking at only 800 Ω x cm2) when compared to freshly derived BMECs. A second passaging of post thawed BMECs exhibited even lower TEER values (peaking at only 200-300 Ω x cm2) (Figure 9) and showed frayed and/or freckled patterns of the tight junction formation (Figure 10). Western blot analysis48 revealed that iPSCs primarily expressed an endodermal marker (SOX17)49 and some tight junction markers (OCLN), but not other BMEC related markers (TJP1, CLDN5, and SLC2A1) (Figure 11). BMECs primarily expressed endothelial related markers (TJP1, CLDN5, OCLN and SLC2A1), with low levels of the endodermal marker, SOX17.

Figure 7: Bright Field Images of Expanded and Cryopreserved iPSC-derived BMECs. A) Day 10 (4 day after initial sub-culture) cells reached maximum confluency. B) Day 11, 24 hours after passaging onto COL4/FN coated 24-wells plate. C) Day12, 48 hours after passaging onto COL4/FN coated 24-wells plate; peak TEER values was observed and ICC was performed. D) 48 hours post-thawed iPSC-derived BMECs on COL4/FN coated 6-wells plate; cells were previously cryopreserved at 1.2 x 102 cells/mL. E) 24 hours after post-thawed iPSC-derived BMECs were passaged onto COL4/FN coated 6-wells plate. F) 48 hours after post-thawed iPSC-derived BMECs were passaged. Please click here to view a larger version of this figure.
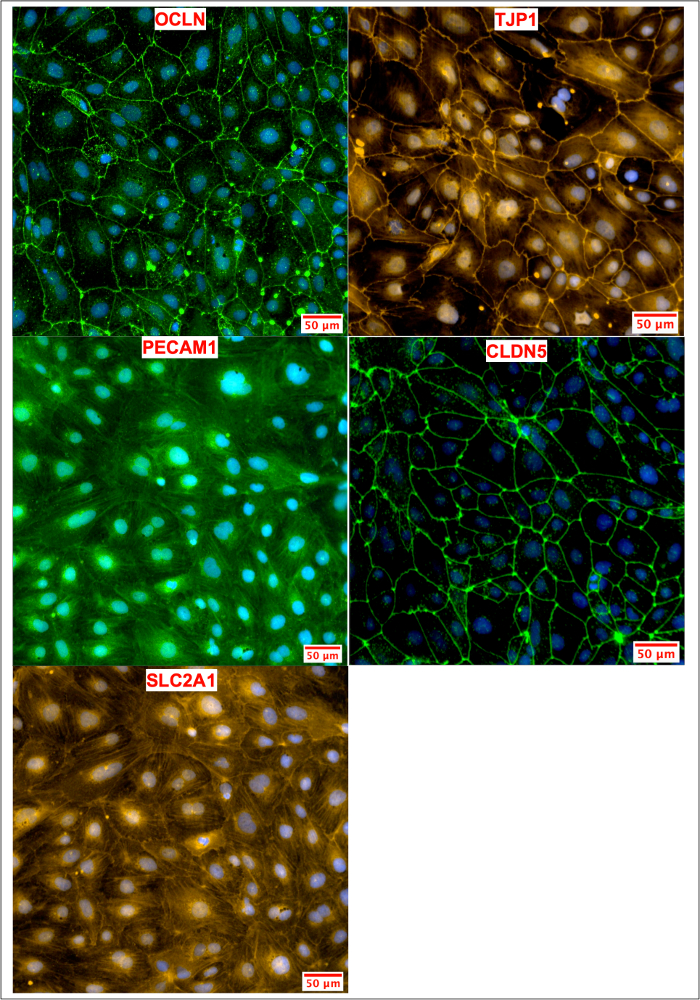
Figure 8: ICC of iPSC-Derived BMECs after Passaging. BMECs were passaged and maintained on COL4/FN matrix until day 12, when TEER values peaked. BMECs on day 12 were stained for tight junction (OCLN, TJP1, CLDN5), influx transporter (SLC2A1) and adherens junction (PECAM1) proteins. The expression pattern and localization resemble those observed in conditions where passaging was not performed, as shown in Figure 4. Please click here to view a larger version of this figure.
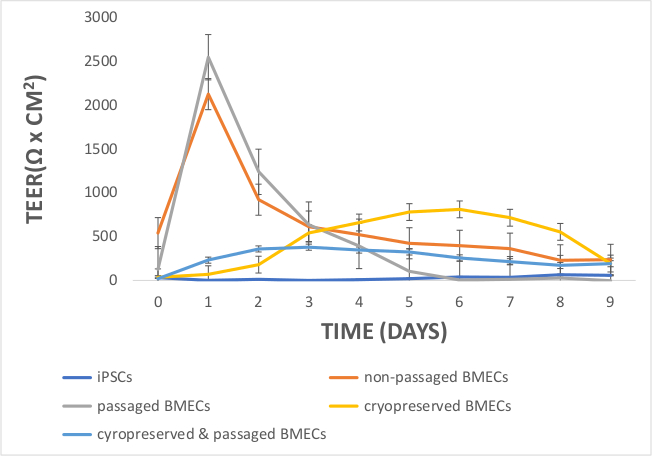
Figure 9: Comparing TEER Measurements in iPSCs, non-passaged BMECs, passaged BMECs, cryopreserved BMECs, and cryopreserved & passaged BMECs. On day 1, TEER values peaked for non-passaged and passaged BMECs, but not iPSCs or cryopreserved BMECs. Cryopreserved BMECs had moderate TEER values between day 3 and 7, with even lower TEER values for the cryopreserved & passaged BMECs. iPSCs did not demonstrate any measurable TEER values between day 0 and 9. TEER measurements were obtained in technical (3 measurements per well) and biological replicates (3 per cell line). The technical average value from a blank well was subtracted from the raw TEER values. These values were averaged for each day and multiplied by 1.12 cm2 (surface area of the 12-transwell insert). Error bars represent standard error. Please click here to view a larger version of this figure.
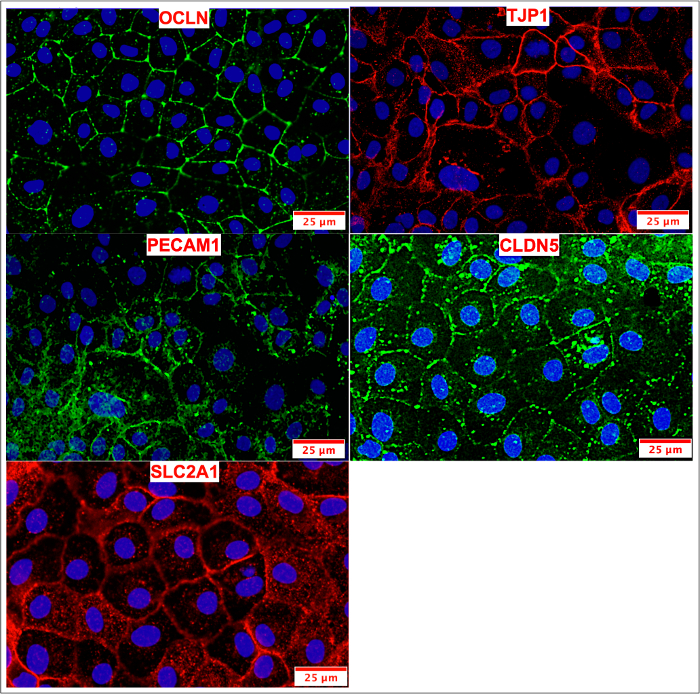
Figure 10: ICC of cryopreserved & passaged BMECs. BMECs were passaged and maintained on COL4/FN until peak TEER values were observed. BMECs were stained for tight junction (OCLN, TJP1, CLDN5), influx transporter (SLC2A1) and adherens junction (PECAM1) proteins. The expression pattern of tight junction markers appeared frayed and/or freckled when compared to non-passaged BMECs (Figure 4) and passaged BMECs (Figure 8). Please click here to view a larger version of this figure.
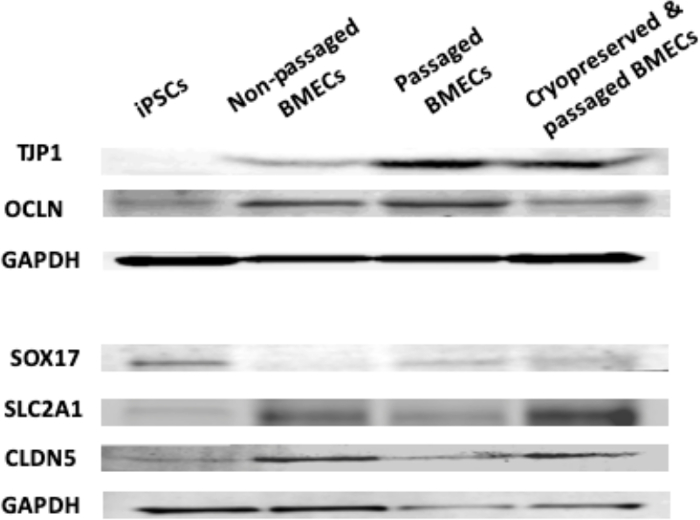
Figure 11: Western blot analysis of iPSCs, non-passaged BMECs, passaged BMECs, and cryopreserved & passaged BMECs. Western blots showing levels of TJP1, OCLN, SOX17, SLC2A1, CLDN5, and loading control (GAPDH). Please click here to view a larger version of this figure.
