To demonstrate the feasibility of the XistΔ:Mecp2/Xist:Mecp2-Gfp mouse model for Xi reactivation studies, XCIF inhibitor-mediated reactivation of Xi-linked Mecp2-Gfp was tested in mouse embryonic fibroblasts (MEFs). Female MEFs were isolated from day 15.5 XistΔ:Mecp2/Xist:Mecp2-Gfp embryos as described in section 3 (Figure 1A). The genotypes of female XistΔ:Mecp2/Xist:Mecp2-Gfp MEFs were confirmed by genotyping-PCR, as described previously19 (Figure 1B), and FACS-based assay (Figure 1C). MEFs were treated with either DMSO or the two drugs LDN193189 and GSK650394 (0.5 μM and 2.5 μM, respectively) for 7 days. Following drug treatment, the expression of Mecp2-Gfp was monitored by qRT-PCR. As shown in Figure 1D, the drug treatment, but not DMSO, reactivated expression of Xi-Mecp2-Gfp. Next, quantitative immunofluorescence was carried out to determine the extent of Xi-Mecp2-Gfp expression in individual MEFs, as described in step 5.3.2. Signal from negative-control MEFs isolated from male embryos (Mecp2/Y) was set as the background, and ~66% of positive control Mecp2-Gfp/Mecp2 MEFs had a nuclear GFP signal. As expected, XistΔ:Mecp2/Xist:Mecp2-Gfp MEFs treated with DMSO had a very low level of nuclear GFP (~3%). By contrast, ~31% of drug-treated XistΔ:Mecp2/Xist:Mecp2 MEFs were positive for nuclear GFP (Figure 1E). Together, these results demonstrate that XCIF inhibitors reactivate Xi-linked Mecp2 in MEFs, but the extent of Xi reactivation varies in the cell population.
To assess the feasibility of pharmacological Xi reactivation-based approach in vivo, whether drug treatment reactivates Xi-linked Mecp2 in the brain of XistΔ:Mecp2/Xist:Mecp2-Gfp female mice was investigated. 10 μL of vehicle or 10 μL of XCIF inhibitors (1.5 mM LDN193189 and 1.6 mM GSK650394) was administered in opposite brain hemispheres of 4-week-old Xi-Mecp2-Gfp female mice by intracerebroventricular injection using stereotactic surgical procedures (Figure 2A,B). The procedure was repeated every second day (Figure 2C), and three weeks later, animals were euthanized, and brains were isolated. One subset was used for qRT-PCR and another was analyzed by immunohistochemistry. The expression of wild-type Mecp2 and Xi-Mecp2-Gfp in the vehicle- and drug-infused hemispheres was determined by qRT-PCR (sequences of primers listed in Table 1). As shown in Figure 2D, drug treatment reactivated Xi-Mecp2-Gfp in ~30% of cells in the drug-infused brain hemisphere, whereas Xi- Mecp2-Gfp was not detected in the vehicle-infused hemisphere. A large number of MAP2, a neuronal marker, positive neurons were also GFP positive (~45%) in the drug-treated hemisphere, indicative of Xi-Mecp2-Gfp expression. Approximately 20% of MAP2 negative brain cells expressed GFP, confirming Xi-Mecp2-Gfp reactivation in non-neuronal cells (Figure 2E).
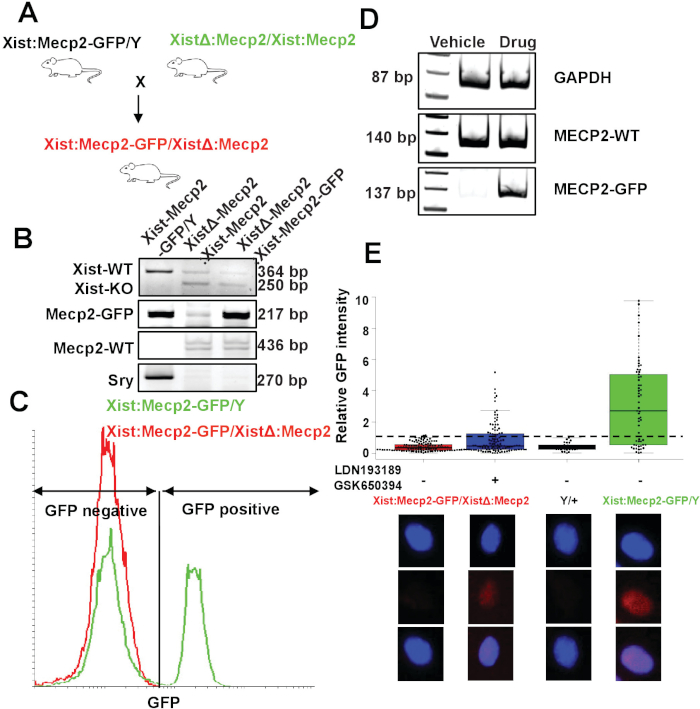
Figure 1: Generation and validation Xi-Mecp2 mouse model. (A) Schematic of the breeding strategy for generating XistΔ:Mecp2/Xist:Mecp2-Gfp mice. (B) PCR genotyping of Xist:Mecp2-Gfp/Y, XistΔ:Mecp2/Xist:Mecp2 and XistΔ:Mecp2/Xist:Mecp2-Gfp mice. Mice were monitored for the presence of Mecp2-Gfp, Mecp2 and sex-determining region Y (SRY). (C) Flow cytometry analysis of nuclei isolated from the mouse cortex. Mecp2/Mecp2-Gfp mouse cortex show ~50% of GFP-positive nuclei while XistΔ:Mecp2/Xist:Mecp2-Gfp shows no GFP-positive nuclei. (D) qRT-PCR analysis monitoring the expression of Mecp2-Gfp and wild-type Mecp2 transcripts in female XistΔ:Mecp2/Xist:Mecp2-Gfp MEFs following treatment with DMSO or drug (LDN193189 and GSK650394). Gapdh was monitored as a loading control. (E) Quantitative immunofluorescence monitoring GFP intensity in female XistΔ:Mecp2/Xist:Mecp2-Gfp MEFs following treatment with DMSO or the drugs LDN193189 and GSK650394. MEFs isolated from Mecp2/Y or Mecp2/Mecp2-Gfp mice were used as negative and positive controls, respectively. Each dot represents an MEF, and the dashed line indicates the maximum background signal obtained in Mecp2/Y, which was set to 1. Lower panel shows representative pictures of nuclei. This figure has been modified from Przanowski et al.16. Please click here to view a larger version of this figure.
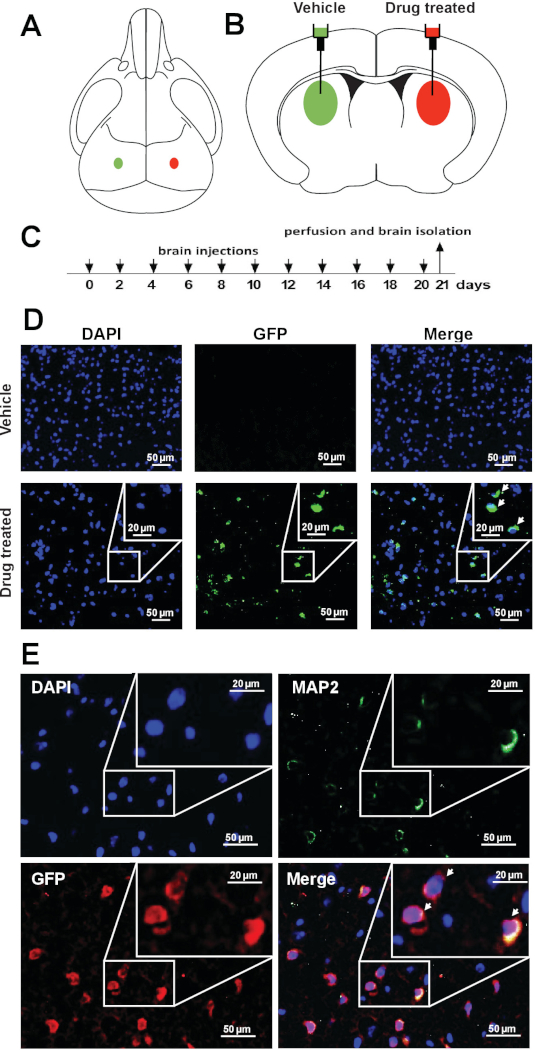
Figure 2: Pharmacological reactivation of X-linked Mecp2 in cerebral cortical neurons of living mice. (A) Schematic of a mouse skull and (B) brain showing the site of injection for vehicle or drug in the left or right hemispheres of the brain. (C) Schematic of the drug regimen. (D) Representative immunofluorescence images showing endogenous GFP signal (green) in coronal brain sections from vehicle- or drug-treated hemispheres. DAPI staining is shown in blue. (E) Representative immunofluorescence images of the coronal brain sections monitoring the expression of GFP (anti-GFP; red) and MAP2 (green) in drug-treated hemisphere. DAPI staining is shown in blue. This figure has been modified from Przanowski et al.16. Please click here to view a larger version of this figure.
| Reverse primer (5’ -> 3’) | Anealing temperature/PCR product length |
| GGCATGGACTGTGGTCATGAG | 60 °C/87 bp |
| GCTGAACTTGTGGCCGTTTA | 62 °C/137 bp |
| TGTCAGAGCCCTACCCATAAG | 62 °C/140 bp |
| GCACAACCCCGCAAATGCTA | 62 °C/364 bp |
| GCACAACCCCGCAAATGCTA | 62 °C/250 bp |
| AATTGCCCTAACGAGCACAC | 62 °C/436 bp |
| GAACTTCAGGGTCAGCTTGC | 62 °C/217 bp |
| CTCCTCTGTGACACTTTAGCCCTCCGA | 66 °C/270 bp |
Table 1: List of primers used for genotyping and quantitative real-time RT-PCR.
