Following the earlier dissection of the ACM and CAM at E5.5 to expose the cranial region of the developing embryo, a series of lacerations that spanned the E7 central cornea was made in ovo (Figure 1). An ideal wound to study cornea regeneration occurs following three lacerations, each made in the same location of the cornea. The first laceration traverses the corneal epithelium, while the second and third lacerations penetrate the underlying basement membrane and anterior stroma, respectively. To achieve an ideal wound, it is crucial to use a sharp micro-dissection knife (see Table of Materials) and apply the correct amount of pressure as the laceration is made (Figure 2, see ideal wound). Applying too little pressure will result in a shallow wound that tears the corneal epithelium without sufficiently penetrating the anterior stroma (Figure 2, see shallow wound). Yet, applying too much pressure results in a full extent wound penetrating the entire stroma and exposing the aqueous humor to the external environment (Figure 2, see full extent wound).
Carrying out the proper lacerating incisions produces an ideal wound (Figure 2) that initially enlarges (0-3 days post wounding)8 (Figure 3). It has been postulated that the phase of wound enlargement that occurs by wounding E7 chicken corneas is related to the rapid expansion of eye size at this embryonic stage8. Embryonic chicken eyes grow at a significantly faster rate from E4 to E10 as compared to the eye growth from E10 to hatching. These early rapid phases of eye growth are attributed to elevated intraocular pressure (IOP)-dependent growth13. Therefore, it is likely that the rapid growth rate of the eye coupled with the elevated IOP promotes wound retraction during the early phases of the healing process (0-3 days post wounding), which is unique to the embryonic cornea wound healing progression. After that, re-epithelialization and new tissue formation occur (4-9 days post wounding) to ultimately close the wound in a scar-free fashion by 11 days post wounding8 (Figure 3A).
Further analysis of wound depth and regeneration was possible by staining cross-sections with a laminin antibody that marks the laminin-rich basement membrane and counter-staining the sections with the nuclear marker DAPI, which reveals the extent of the wound through the corneal epithelium8. Recently wounded corneas (0 dpw) and those that are early in the regeneration process (3 dpw) showed that the wound penetrated the epithelial layer and basement membrane, as evidenced by the break-in staining of the nuclear marker DAPI within the corneal epithelium and the absence of laminin antibody staining, which marks the laminin-rich basement membrane between the corneal epithelium and underlying stroma8 (Figure 3B). However, cross-sections through 11 dpw corneas stained with DAPI and laminin antibody revealed a completely healed cornea that had been re-epithelialized and contained a continuous laminin-rich basement membrane at the site of the regenerated wound8 (Figure 3B).
Following the corneal incision, detailed characterization of the wound healing process was accomplished by performing immunohistochemistry on sectioned, wounded corneal tissues. The extracellular matrix proteins fibronectin and tenascin are associated with epithelial and keratocyte cell migration into healing adult corneal wounds14,15. Spatiotemporal localization of the extracellular matrix proteins, fibronectin and tenascin is apparent within the healing wound and was found to be elevated at timepoints corresponding to corneal re-epithelialization (5 days post wounding)8 (Figure 4). Such analysis suggests the importance of fibronectin and tenascin to wound closure and, specifically, their involvement in epithelial cell migration and survival, consistent with such functions in adult corneal wounds16,17.
Commencing at E8-E9, the cornea becomes densely innervated by trigeminal sensory nerve fibers that emanate from a pericorneal nerve ring and traverse through the anterior stroma as they project toward the cornea's center and the corneal epithelium by E1218,19,20. Since corneal wounds in this model are made at E7, shortly before nerve projection into the cornea, this model further investigates corneal nerves as they navigate a healing cornea following insult. By using whole-mount immunohistochemistry to trace corneal nerves with anti-β neural tubulin (Tuj1) antibodies21, it is apparent that nerves are temporarily inhibited from the healing corneal tissue that directly juxtaposes the wounded, central cornea (5 days post wounding)8 (Figure 5A,B). Despite earlier inhibition, corneal nerves eventually innervate the fully healed corneal tissue (11 days post wounding) to similar density levels and in similar patterns to stage-matched, non-wounded controls (E18C) (Figure 5C,D).
Strikingly, fully re-epithelialized corneal tissues that have healed in a nonfibrotic, scarless fashion display a complete recapitulation of the normal collagen tissue architecture. As evidenced by second-generation harmonic imaging22,23, bundles of collagen fibers throughout varying depths of the central cornea wound area are arranged orthogonally, matching the native macrostructure of non-wounded central corneal tissue9 (Figure 6).
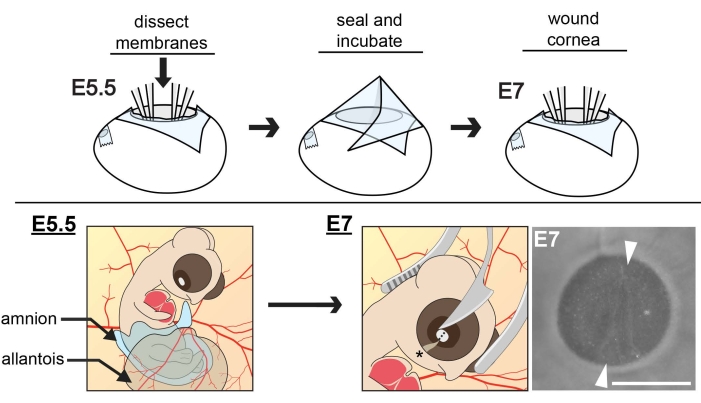
Figure 1: Schematic of in ovo extraembryonic membrane dissections and corneal wounding. At E5.5, the cranial region of the embryo is exposed by dissecting the ACM and CAM membranes and positioning the amnion and allantois away from the developing eye. Eggs are sealed and incubated to E7 when the central cornea is wounded, using curved forceps as a cradle for the embryo head as the tip of a microsurgical knife makes an incision in the central cornea. The wound is oriented parallel to the choroid fissure (asterisk). To ensure the depth of the incision reaches the anterior stroma, three concurrent cuts need to be made with the knife, each in the same relative position, one over the other. Scale bar = 1 mm. The figure is adapted with permission from references8,11. Please click here to view a larger version of this figure.
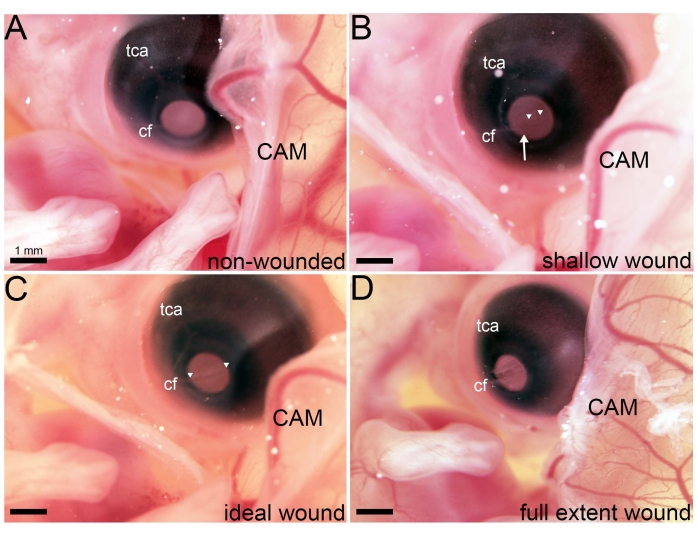
Figure 2: Variations in wounds generated in ovo. (A) Following membrane dissections at E5.5, the right eye is accessible in ovo. (B–D) Images taken of an in ovo embryo immediately following lacerations of varying degrees that span the cornea's extent and are in line with the choroid fissure (cf). (B) Following three lacerations wherein weak pressure was applied, a shallow wound is visible. The arrowhead marks a site in the cornea where the epithelium has been sheared but the anterior stroma has not been penetrated. Arrowheads denote a small cornea region where the anterior stroma has been penetrated. (C) Following three lacerations wherein an ideal amount of pressure was applied, an ideal wound is visible. Arrowheads denote a wound spanning the entire extent of the cornea where the anterior stroma has been penetrated. (D) Following three lacerations wherein excessive pressure was applied, a full extent wound is visible, and the aqueous humor has become exposed to the external environment. Abbreviations: tca, temporal ciliary artery; cf, choroid fissure; CAM, chorioallantoic membrane. Please click here to view a larger version of this figure.
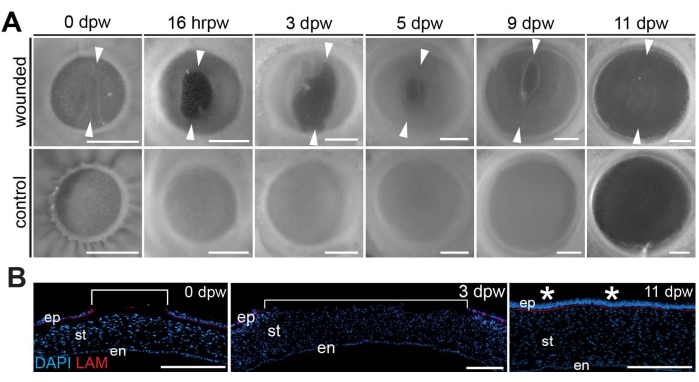
Figure 3: Wound healing progression. (A) Progression of healing in wounded corneas compared to stage-matched controls is shown from the time of wounding (0 dpw) and 16 h post-wounding (hrpw) through 3-11 days post wounding (dpw). Arrowheads delineate the dorsal- and ventral-most borders of the wound, indicating a period of wound expansion (0-3 dpw) followed by progressive wound closure (5-11 dpw). (B) Sectioned DAPI (blue)- and laminin (red)-stained wounded corneas at 0, 3, and 11 dpw. Brackets show the extent of the wounded region, which reveals wound expansion by 3 dpw and full repair of the re-epithelialized cornea by E11. Asterisks denote the healed region of the cornea. The scale bar is (A) 1 mm, (B) 100 µm. The figure is adapted with permission from reference8. Please click here to view a larger version of this figure.
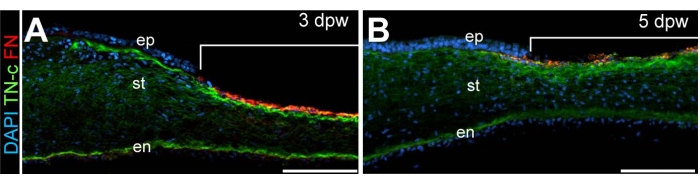
Figure 4: Histological analysis of wounded corneas. Cross-sections through (A) 3 dpw and (B) 5 dpw. DAPI-stained (blue) wounded corneas reveal the localization of fibronectin (FN, red) and Tenascin-C (TN-C, green). Brackets in (A) and (B) denote the wounded region. Scale bar: 100 µm. Abbreviations: ep, epithelium; st, stroma; en, endothelium. The figure is adapted with permission from reference8. Please click here to view a larger version of this figure.
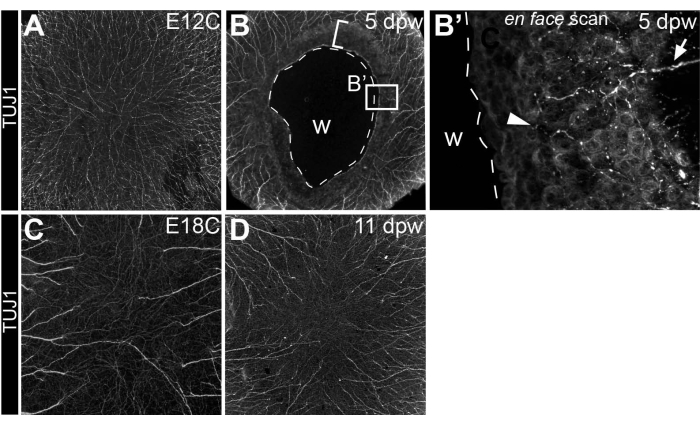
Figure 5: Innervation of wounded corneas. (A–D) Visualization of the corneal nerves following anti-β neural tubulin (Tuj1) whole-mount immunostaining in (B) 5 dpw and (D) 11 dpw corneas, as well as (A) stage-matched E12 (E12C, stage-matched for 5 dpw) and (C) E18 controls (E18C, stage-matched for 11 dpw). (B) The broken line in the 5 dpw cornea denotes the extent of the wound. The bracketed area directly adjacent to the wound denotes the cornea area that is temporarily repulsive to nerves and actively undergoing re-epithelialization. (B') Optical scan through the healing corneal tissue directly adjacent to the open wound reveals a rare stromal nerve bundle extending into the tissue (arrow) and epithelial nerve leashes (arrowhead). (C,D) Fully regenerated corneas at 11 dpw display similar innervation patterns and comparable nerve densities to stage-matched controls. Abbreviation: w, wound. The figure is adapted with permission from reference8. Please click here to view a larger version of this figure.
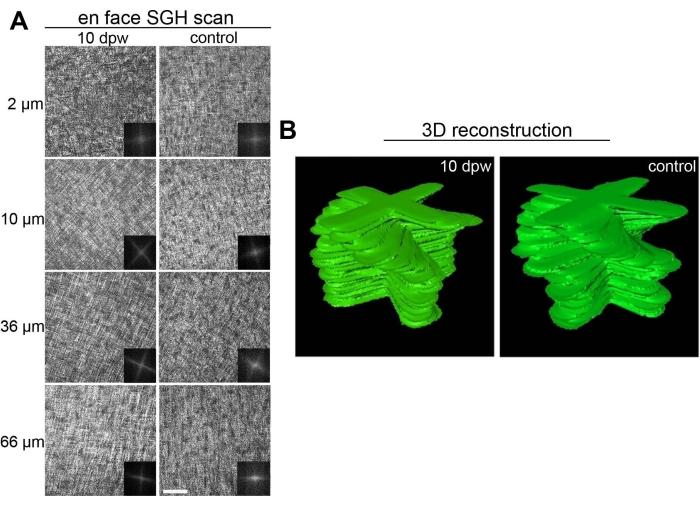
Figure 6: Collagen ultrastructure in healed embryonic corneas. (A) En face scan of fully healed 10 dpw corneas and stage-matched controls using second-generation harmonic imaging (SGH). Scan depths range from 2-66 µm from the anterior surface of the cornea (0 µm is the most anterior stroma) and are listed to the left of the respective images. Insets for each image correspond to Fast Fourier transform analysis of the central wound area for that particular scan depth. (B) Manually segmented stacks of two-dimensional Fast Fourier transform analysis, representing collagen organization within the wounded and stage-matched control cornea. Scale bar = 50 µm. The figure is adapted with permission from reference9. Please click here to view a larger version of this figure.
