Procedures involving mice were approved by the Institutional Animal Care and Use Committee (IACUC) at Brigham Young University. Male and female (n = 8) C57BL/6 mice aged to at least P21 were used in the following experiments.
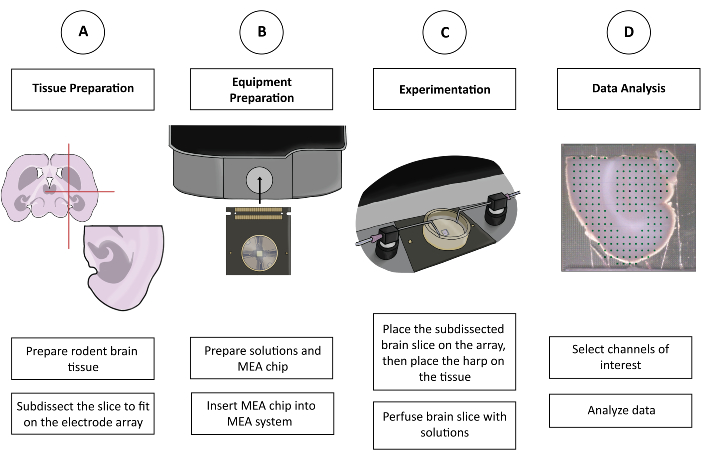
Figure 1: Schematic figure of CMOS-HD-MEA experimentation. (A) The brain slice is prepared by one's preferred cutting method and sub-dissected to fit on the MEA. (B) Prepare the solutions and the CMOS-HD-MEA chip. (C) The sub-dissected brain slice is placed on the electrode array and bathed in the appropriate solutions. (D) Relevant channels are selected from the collected data. Data is then prepared for analysis in the preferred program of the user. Please click here to view a larger version of this figure.
| Name | Concentration (mM) | g/L | ||
| Sodium Chloride (NaCl) | 126 | 7.36 | ||
| Potassium Chloride (KCl) | 3.5 | 0.261 | ||
| Dihydrogen Sodium Phosphate (NaH2PO4) | 1.26 | 0.151 | ||
| Sodium Bicarbonate (NaHCO3) | 26 | 2.18 | ||
| Glucose (C6H12O6) | 10 | 1.80 | ||
| Magnesium Chloride (MgCl2) | 1 (from 1 M stock) | 1 mL | ||
| Calcium Chloride (CaCl2) | 2 (from 1 M stock) | 2 mL | ||
Table 1: aCSF solution.
1. Preparing solutions
- Prepare the experimental solutions.
- Prepare 1 L of artificial cerebrospinal fluid (aCSF) (see Table 1 for details).
- Prepare 1 L of pro-convulsant solution.
NOTE: Solutions used to generate seizure-like activity in this protocol utilized either aCSF with 100 µM 4-Aminopyridine (4-AP) or aCSF free of magnesium ions. - Carbogenate all solutions for at least 10 min before use using porous stones.
- Obtain a beaker for discarding solutions.
- Place 1 L of aCSF, 1 L of pro-convulsant solution, and the discard beaker on a surface close to the perfusion system.
- Preparing the cutting and holding chamber solutions
- Prepare 0.5 L of aCSF, omitting CaCl2 and adding 3 mM MgCl2 (as opposed to 1 mM) to create a unique cutting solution. Retain this solution for use in rodent brain slice preparation by placing it near the acute brain slice preparation area.
- Prepare 0.5 L of aCSF and place it in a tissue-holding chamber that will be used to retain slices for use in experimentation. Insert porous stones into the tissue holding chamber and carbogenate the solution for at least 10 min before placing the sub-dissected slices in the chamber.
2. Preparing rodent brain slices
- Prepare all solutions as described above (see section 1). Ensure that all solutions are properly carbogenated during the preparation process.
NOTE: The cutting solution (see section 1.2) is recommended, but other cutting solutions can be used to obtain quality brain slices, such as a sucrose cutting solution30. - Use a vibrating microtome to make 350 µm rodent brain slices. Protocols for using these instruments are listed in the references30,31,32,33.
- Prepare the rodent brain slices to fit on the recording electrode area of the MEA chip (see Figure 2). Use a size 10 surgical blade to sub-dissect the slices, as shown in Figure 1A and Figure 2A , by gently rolling the blade back and forth on the brain slice. Perform the sub-dissection in the vibrating microtome cutting chamber. Use caution not to damage the sub-dissected brain slice.
- Place the sub-dissected slices in a tissue-holding chamber filled with aCSF. Ensure that the chamber has been carbogenated for at least 10 min before adding the sub-dissected slices.
3. Equipment preparation
- Preparing the MEA chips and system (Figure 2 and Figure 3)
NOTE: Hard materials like plastics and metals can easily damage the MEA chip if they forcefully contact the bottom of the chip well. When using pipettes to transfer the solution into or out of the chips, do not contact the bottom of the chip well with the pipette, especially the reference electrodes on the sides of the chip well and the recording electrodes in the center of the chip well (Figure 2C). Instead, add the solution by hovering over the bottom of the chip well or by contacting or approximating the plastic edges of the chip well. To pull out all solution easily, tilt the chip slightly to allow the solution to pool to one side of the chip well and remove it from the top of the pooled solution, or use an anti-static wipe to dab lightly at areas that still have the solution. Do not transport the chip by holding the chip well or the chip pins (Figure 2C). The chip well can hold approximately 4 mL of solution. For subsequent steps, fill the chip well with approximately 2 mL of solution if not otherwise specified.- Designate transfer pipettes for various tasks before beginning chip preparation. Label one transfer pipette for ethanol, one for waste, another for aCSF, and others for any remaining solutions to prevent unintentional mixing.
- Fill the well of the MEA chip with 190-proof ethanol so that the bottom of the chip well is completely covered (Figure 3). Let the ethanol sit for 30-60 s, then remove it with a discard pipette.
- Fill the well of the MEA chip with aCSF and remove it with a discard pipette to rinse residual ethanol out of the chip well. Add and remove aCSF from the chip well three times, using the previously designated waste and aCSF pipettes. After washing the chip well three times, add aCSF and let it stand for at least 30 s.
NOTE: The recording electrodes exhibit the least amount of noise from hydrophobic interactions when aCSF remains in the chamber for at least 45 min after rinsing with ethanol and aCSF. - Before docking the MEA chip, wet an anti-static wipe with 190-proof ethanol and use it to wipe the chip's pins (Figure 2C).
- Gently slide the MEA chip into the MEA platform and engage the docking mechanism to lock the chip into place.
- Check the recording and reference electrodes for bubbles (Figure 2C). If bubbles are present, take a clean paintbrush and lightly sweep over the electrodes to remove them.
- Check the chip for noise using the CMOS-HD-MEA software34 and visually scan the false color map for bubbles, non-biological oscillations, or spikes caused by electrical interference. Ground the MEA system appropriately to negate any encountered noise.
NOTE: The grounding setup will depend on the recording environment. For the experiments in this protocol, the MEA platform and the perfusion system were grounded.
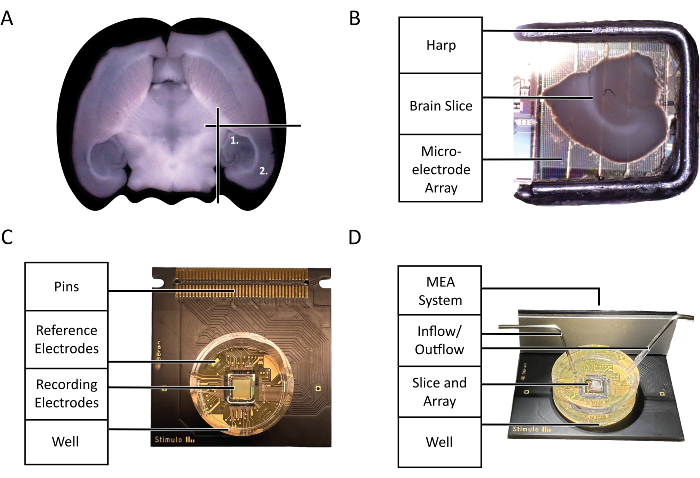
Figure 2: Configuration and technology diagrams. (A) Diagram of the selection of acute mouse brain slices used in the experiment highlighted by this protocol. (1) Hippocampal region (2) Neocortex region. (B) The proper placement of an acute mouse brain slice and harp on the microelectrode Array (MEA). (C) The anatomy of a 3Brain Accura CMOS-HD-MEA chip. (D) The proper configuration of perfusion inlets and outlets. Input should be deep in the chip well, whereas the output should be on the opposite side of the inlet at the top of the chip well to guarantee a constant flow of fresh, oxygenated aCSF. Please click here to view a larger version of this figure.
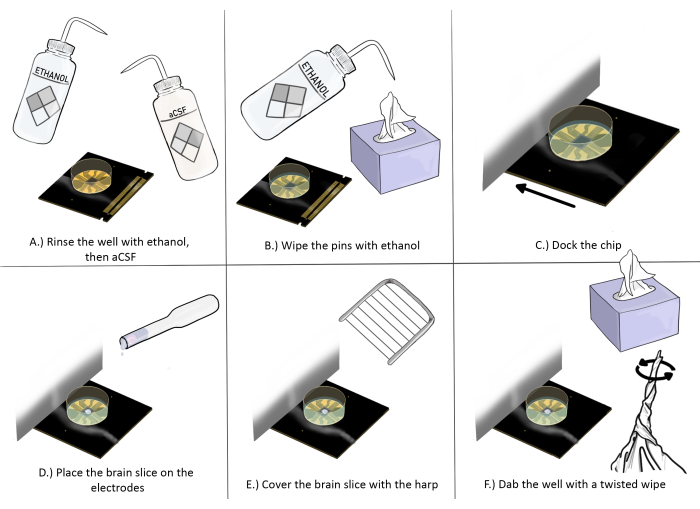
Figure 3: Schematic diagram of chip preparation and placement for brain slice experiments. (A) Rinse the chip well with ethanol once, then aCSF three times. (B) Wipe the pins with ethanol using an anti-static wipe. (C) Dock the chip. (D) Place the brain slice on the electrodes. (E) Place the harp on the brain slice (see Figure 2 for proper placement guidelines). (F) Dab the corner of the recording electrodes' well near the brain slice with a twisted anti-static wipe. Please click here to view a larger version of this figure.
4. Experimentation
- Placing the brain slice
- Place a platinum harp in a weigh boat (or another clean surface) near the MEA platform. Cover the harp with about 3mL of aCSF to reduce its hydrophobic tendencies.
- Use scissors to remove the thin tip portion of a transfer pipette. Cut off a third, about 1.5 in, of the pipette tip.
NOTE: This modified pipette will not constrict or damage the brain slice during collection and transportation. - Collect a brain slice from the slice-holding chamber with the modified pipette. Gently dispense the brain slice and any solution in the pipette into the chip well. To position the slice properly, gently release more aCSF from a transfer pipette to manipulate the position of the brain slice or use a soft paintbrush to create a current in the solution that pushes the brain slice onto recording electrodes. Limit contact with the recording electrodes or brain slice to minimize damage.
- Using forceps, gently place the harp over the brain slice with the threads downwards to press the slice onto the recording electrodes. Avoid contacting the electrode array with the harp. Orient the harp so that the side without a frame faces toward the inflow needle, and the frame of the harp does not contact any of the recording electrodes (Figure 2C, D).
- Take a discard pipette and remove excess aCSF. Take an anti-static wipe, twist a corner to create a tip, and use it to soak up the remaining aCSF surrounding the recording electrodes without touching the recording electrodes, brain slice, or harp (Figure 2).
- Using a designated aCSF pipette, quickly add enough carbogenated aCSF to cover the brain slice, about 2 mL.
- Repeat step 4.1.5. and 4.1.6. two more times.
- Fill the well with carbogenated aCSF until the well is roughly ¾ full, about 3 mL.
- Take a picture of the brain slice on the MEA chip with a microscope or camera. Ensure that the photo has a high enough resolution to see the borders of the recording electrode array and the anatomy of the brain slice.
- Running the experiment
- Operating the perfusion system
- Place the inflow and outflow tubes into the beaker filled with aCSF and the inflow and outflow needles into the chip well. Place the inflow needle close to the bottom of the chip well just outside the recording electrodes. Place the outflow needle close to the top of the chip well towards the edge so that the liquid rises almost to the brim of the chip well, about 4 mL, and the slice is submerged throughout the experiment (Figure 2D).
- Set the perfusion inflow to 5 mL/min and the perfusion outflow to 7 mL/min.
NOTE: It is recommended that the outflow rate exceeds the inflow rate to avoid solution overflow out of the chip well and to create a solution current over the brain slice. - Turn on the inflow and outflow. Remove the inflow needle from the chip well until the needle begins to output solution instead of air. Then, place the needle back to its position inside the chip well as described in step 4.2.1.1.
- Use a solution heater to keep the solution at or near physiological temperature, around 34-37 °C.
- Let the aCSF perfuse over the brain slice for 10 min. This will allow the slice to adjust to the recording environment.
- After 10 min has elapsed, move the outflow tube to the discard beaker. Then, move the inflow tube to the beaker containing the pro-convulsant solution. Allow the non-convulsant aCSF to be flushed out of the perfusion system into the discard beaker for 10 min.
- Transfer the outflow tube into the beaker containing the pro-convulsant solution.
- Let the pro-convulsant solution cycle until the experiment finishes.
- If the slice recording duration exceeds 2 h, consider preparing another pro-convulsant solution to offset the glucose consumption of the active brain slice.
- Exchanging brain slices
- Turn off the inflow. Turn off the outflow 10-15 s afterwards.
- Use forceps to remove the harp and place it onto a weigh boat or other surface.
- Use a modified pipette to extract and discard the brain slice. Do not touch the reference electrodes or the recording electrode array (Figure 2C).
- Place the inflow tube into the beaker containing the non-convulsant aCSF solution. Place the outflow tube in the discard beaker. Leave the inflow and outflow needles in the chip well. Run the perfusion system for 10 min to remove all residual pro-convulsant solution from the chip well and perfusion system. Begin the experimentation process again with a new brain slice (beginning at step 4.1).
- Operating the perfusion system
- Finishing the experiment
- Cleaning the rig
- Remove the harp, brain slice, and solution from the well, respectively.
- Undock the MEA chip, place it onto a clean surface, and fill the chip well with deionized water.
- Discard the deionized water to clear any salts left from the aCSF solutions.
- Using a transfer pipette, fill the chip well with a detergent solution. Hover the tip of the transfer pipette in the chip well and point it at the recording electrodes. Quickly and repeatedly squeeze and release detergent into the transfer pipette to vigorously wash detergent over the recording electrodes for 1 min. Let the detergent rest for 5-10 min.
- Remove the detergent, then rinse the chip well 4-6 times with deionized water to remove the detergent solution and any remaining solutes.
- Remove all the water from the chip well, then place an anti-static wipe over the MEA chip and leave overnight to allow the chip well to dry.
- Place a weigh boat or another watertight plate onto the MEA recording platform where the MEA chip previously rested. Place the harp in the center of the plate and place the inflow and outflow needles on either side of the harp.
- Clean the perfusion system and harp by sending 50 mL of water, 50 mL of 140-proof ethanol, and 200 mL of water, respectively, through the tubing into the discard beaker.
NOTE: At this time, perfusion speeds can be increased to decrease cleaning time, but the outflow speed must always exceed the inflow speed to avoid flooding. - Clean the carbogen stones by placing them in a beaker of distilled water and sending the carbogen through the tubing for 5 min. Set them out to rest on a dry, clean surface overnight and cover them to avoid dust accumulation.
- Cleaning the rig
5. Data analysis
NOTE: There are a variety of analysis packages used for analyzing electrophysiological data produced by CMOS-HD-MEAs, including BrainWave from 3Brain, Yet Another Spike Sorter (YASS), and custom Python tools34,35,36,37. We extracted data from the BrainWave data file format for use in the Xenon LFP Analysis platform to generate the data presented in Figure 4 and Figure 5. Custom Matlab code was used to analyze the data in Figure 6. Protocols for the Xenon LFP Analysis platform are publicly available10. The following protocol steps are specific to recordings made with Brainwave 438; for other systems, refer to supporting documentation related to those systems34,35,36,37. An overview of the analysis steps taken to produce the data with this protocol is provided below. For complete details of how to export, visualize, and analyze data, including tutorial videos and all relevant code files, see39.
- Export the recording file for analysis.
- Create a folder for the data recording file(s) that will be downsampled.
- Crop the image taken during step 4.1.9 to include only the recording array with the brain slices.
- Open the ExportToHDF5-ChannelSelection.py file (this code file can be accessed on GitHub under xenon-lfp-analysis/code-files/3Brain-processing40) and copy and paste the URL listed at the top of the window into an internet browser.
- Input the file path for the data recording file and upload the cropped image of the array associated with the recording.
- Under Select Channels for Export, use the lasso select tool to select the hippocampal and neocortical regions shown on the cropped image of the array.
- Set both the number of rows to skip and the number of columns to skip to 2.
NOTE: Recording files are often large; if appropriate for the desired analysis, the exported data can be downsampled. Options exist to downsample spatially (by skipping rows and columns of recording electrodes) and temporally (by downsampling to a lower sampling frequency). In the analysis, every 2 rows and 2 columns are skipped; this allows downsampling of the data spatially without being biased to certain channels. Each channel is far enough apart that the differences in activity could be significant between them; therefore, no channel averaging is performed. When skipping rows and columns of the electrodes, the data from unselected channels are not included when the output file is exported. - Set the downsampling frequency to 300 Hz.
- Click the Export Channels button to generate the channel selection file.
NOTE: The software will automatically save the channel selection file to the folder where the data recording file is saved. - Open the ExportToHDF5-ExtractDownsample.py file (also found on GitHub under xenon-lfp-analysis/code-files/3brain-processing40), paste the file path of the folder containing the channel selection file(s) and the data recording file(s), and push enter on the keyboard to generate the downsampled version of the data recording file.
- Using the analysis platform
- In the computer's command prompt, type run_lfp_analysis and hit enter to open the analysis platform.
- Input the file paths of the image and downsampled recording file and click submit.
- Under Select a Time Range for Analysis, select the entire recording.
- Under Select Channels for Plots, highlight the hippocampus as Group-1 and the neocortex as Group-2.
- Under Channel Raster, set the Threshold to 0.06 mV and the Time Duration to 0.02 s.
- Click Apply Settings and Generate Raster.
- Use the raster plot to explore different channels that show activity patterns of interest.
NOTE: See Figure 4 for examples of high-quality seizure-like activity (Figure 4A-D) versus suboptimal activity (Figure 4E-F).
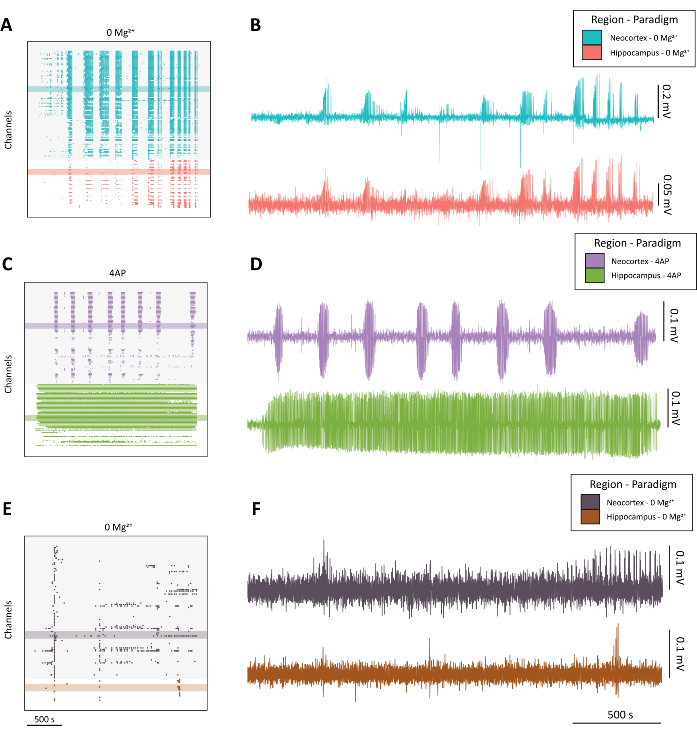
Figure 4: Example of evolving epileptiform activity from the 0 Mg2+ and 4-AP paradigms. (A) Example raster plot from the application of aCSF with 0 Mg2+ over approximately 40 min. (B) Example electrophysiology traces taken from the neocortex (blue) and hippocampus (red) demonstrating epileptiform activity from the 0 Mg2+ paradigm. (C) Example raster plot from the application 100 mM 4-AP over approximately 40 min. (D) Example electrophysiology traces taken from the neocortex (purple) and hippocampus (green) demonstrating epileptiform activity from the application of 4-AP. (E) Example raster plot from the application of aCSF with 0 Mg2+ over approximately 40 minutes showing bursting activity as opposed to seizure-like activity as found in the other representative traces. (F) Example electrophysiology traces taken from the neocortex (dark purple) and hippocampus (rust) demonstrating sub-optimal activity from the 0 Mg2+ paradigm intended for comparison to the quality seizure-like activity found in B and D. Please click here to view a larger version of this figure.
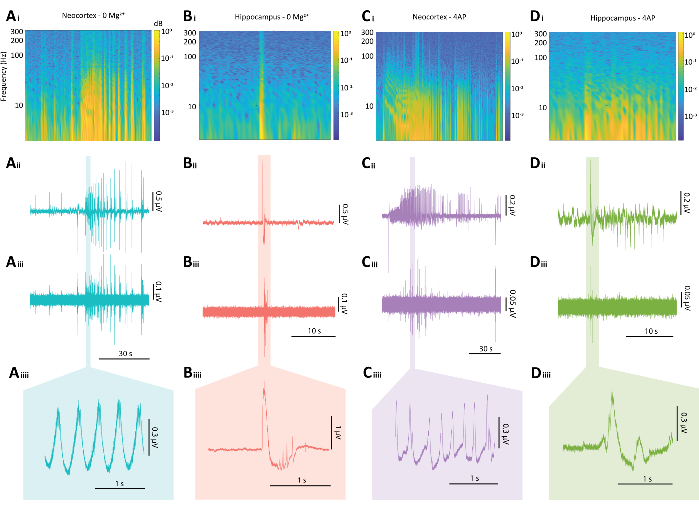
Figure 5: Representative results of epileptiform discharges from both the 0 Mg2+ and 4-AP paradigms. (A) Example plots of a typical neocortical seizure-like event induced by the 0 Mg2+ paradigm including (Ai) a spectrogram from a seizure-like event, (Aii) the associated electrophysiological trace, (Aiii) an 80 Hz high pass filter applied to the trace from Aii, (Aiiii) and a magnified section of the trace from Aii. (B) Example plots of a typical hippocampal epileptiform burst induced by the 0 Mg2+ paradigm including (Bi) a spectrogram of the epileptiform burst, (Bii) the associated electrophysiological trace, (Biii) an 80 Hz high pass filter applied to the trace from Bii, (Biiii) and a magnified section of the trace from Bii (C) Example plots of a typical neocortical seizure-like event induced bythe 4-AP paradigm including (Ci) a spectrogram of epileptiform activity, (Cii) the associated electrophysiological trace, (Ciii) an 80 Hz high pass filter applied to the trace from Cii, (Ciiii) and a magnified section of the trace from Cii (D) Example plots of a hippocampal epileptiform burst under the 4-AP paradigm including (Di) a spectrogram of epileptiform activity, (Dii) the associated electrophysiological trace, (Diii) an 80 Hz high pass filter applied to the trace from Dii, (Diiii) and a magnified section of the trace from Dii. Please click here to view a larger version of this figure.
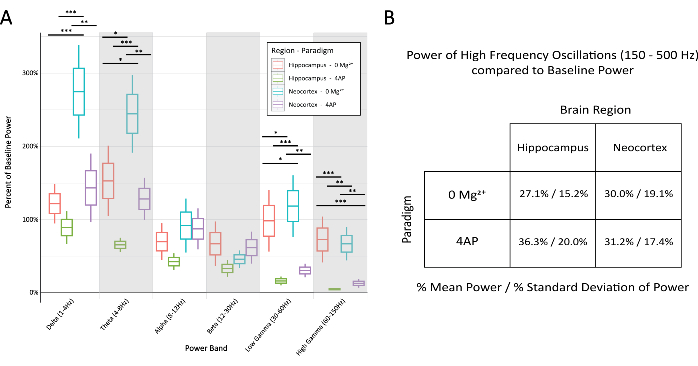
Figure 6: Comparison of percent of baseline power in the various bands across paradigm and brain region during stereotypical epileptiform discharges. (A) The power during epileptiform discharges was significantly different between the paradigms and brain regions for most frequency bands (2-way ANOVA with Tukey test, *P < 0.05, **P < 0.001, ***P < 0.0001). The middle line for each box represents the mean, the borders of the box ±1 standard error of the mean (SEM), and the outermost lines ±2 SEM. (B) Both paradigms and brain regions demonstrated limited power in bands related to high-frequency activity above 150 Hz. Please click here to view a larger version of this figure.
