The aim of this protocol was to highly purify and culture both primary CN3s/CN4s and SMNs long-term to enable comparative analyses of the mechanisms underlying motor neuron disorders (see Figure 1 and Figure 2 for overview).
Once neurons were successfully isolated and grown in culture, nearly pure primary CN3/CN4 and SMN cultures were obtained (Figure 5A,B) and maintained for at least 14 DIV (Figure 4 and Figure 6). The purities of CN3/CN4 and SMN cultures at 2 DIV were 93.5 ± 2.2% and 86.7 ± 4.7%, respectively, when assessed by ICC using the motor neuron marker Islet1 and neuronal marker TUJ1 (Figure 5B). However, these high purities relied heavily on the age of the embryos and on setting appropriate thresholds for GFP gates during FACS (Figure 3). Dissection of embryos at E10.5 is more difficult than dissection at E11.5 due to increased softness and adhesiveness of tissues, resulting in decreased motor neuron yields. However, the purities of E10.5 CN3s/CN4s and SMNs were comparable to those for E11.5 embryos (92.8% and 82.2% at 2 DIV, respectively; data obtained from a single experiment). The purities of CN3s/CN4s and SMNs dramatically decreased when E13.5 embryos were used, even if only the highest GFP-positive population was collected (20.7% and 7.4% at 2 DIV, respectively; data obtained from a single experiment), probably due to the expression of GFP in non-motor neurons (Figure 7). This same tendency also held true for E12.5 cultures, although it was much less dramatic. Therefore, embryos at E12.5 or older are inappropriate for use in the purification of motor neurons using this protocol.
Pure motor neuron cultures are valuable for understanding isolated growth patterns, behaviors, and vulnerabilities of motor neurons. This example demonstrates how these cultures can be used to test motor neuron responses to chemical treatment. To determine if primary CN3s/CN4s and SMNs show differential responses to endoplasmic reticulum (ER) stressors, primary monocultures of CN3s/CN4s and SMNs were obtained using this protocol and treated with varying concentrations of an ER stressor, cyclopiazonic acid (CPA). Neurons were treated with CPA (5, 10, 15, 20, 25, or 30 µM) or vehicle control (DMSO) at 2 DIV and fixed 3 days later for ICC to evaluate survival ratios (Figure 8A). The number of viable neurons in each sample was counted and survival ratios were calculated as the number of viable cells in drug-treated wells divided by the number of viable cells in the wells treated with DMSO. CN3/CN4 monocultures were significantly more resistant to CPA treatment (10−25 µM) as compared to SMN monocultures (Figure 9 and Figure 8B)31.
In conclusion, this protocol allows for the generation of highly purified primary mouse embryonic CN3/CN4 and SMN cultures that provide a powerful and reliable system for the investigation of neuronal behavior.
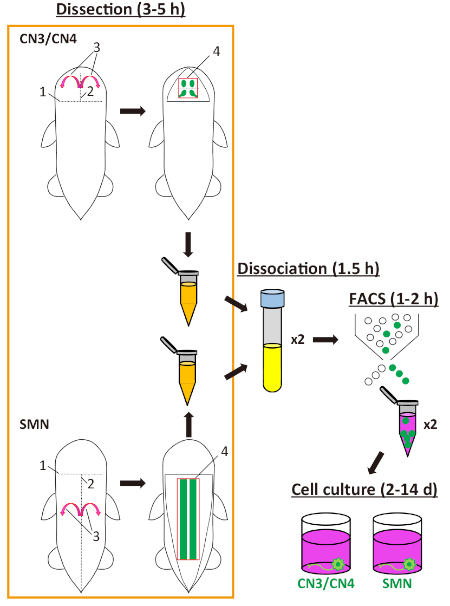
Figure 1: Scheme for preparation of mouse embryonic motor neurons. The schematic illustrates the steps involved in the isolation and culture of mouse embryonic motor neurons and the approximate time in hours or days for each step. The order of the dissection procedure for CN3/CN4 and for SMN are each labelled sequentially 1 through 4. Abbreviations: CN3/CN4 = oculomotor neuron/trochlear neuron; SMN = spinal motor neuron; FACS = fluorescence-activated cell sorting; h = hour; d = day. Please click here to view a larger version of this figure.
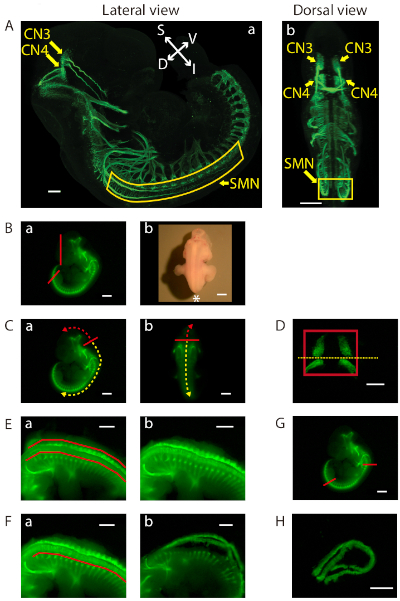
Figure 2: Dissection of the ventral midbrain and the cervical (C1)-lumbar (L2-L3) portion of the ventral spinal cord. (A) Lateral (a) and dorsal (b) views of GFP-positive motor neurons in an E11.5 IslMN:GFP transgenic mouse embryo under fluorescein isothiocyanate (FITC) illumination. A whole mount E11.5 embryo was prepared as previously described32 in order to make the embryo transparent. Subsequently, the embryo was analyzed by immunofluorescence labeling with anti-GFP staining (green). Images were captured under a confocal microscope. Scale bars = 200 µm (lateral view) and 400 µm (dorsal view). Abbreviations: S = superior; I = inferior; V = ventral; D = dorsal. (B-H) Dissection steps highlighted on images of E11.5 ventral midbrain and ventral spinal cord tissues taken with an equipped camera under bright light (Bb) or FITC illumination using a fluorescence dissection stereomicroscope. Scale bars = 200 µm (D), and 1 mm (A−C, E−H). (B) (a) Removal of the face and tail of the embryo by cutting along the red lines. (b) Embryo positioned for dissection. Positioning of the front of the microscope is indicated by an asterisk. (C) Cutting along the solid red line in order to slit open the roof of the fourth ventricle (a) lateral view and (b) dorsal view. Use of this opening to cut along the surface of the embryo dorsal to the brain (trajectory indicated by dashed red arrow). This exposes the tissue containing mesenchyme, CN3, and CN4, which can be lifted out of the cranium. For SMN dissection, insertion of forceps into the same opening between the fourth ventricle and its roof, then cutting toward the caudal side of the embryo (trajectory indicated by dashed yellow arrow). (D) Final view of the ventral midbrain containing bilateral GFP-positive CN3 and CN4 nuclei. The edges of the tissue are highlighted by a red rectangle. Cutting along yellow dotted line to collect CN3 and CN4 nuclei separately, if desired. (E) After opening the rest of the hindbrain and spinal cord, flapping dorsal tissues pinched off above the red lines on both sides with tweezers (a) before, and (b) after. (F) Bilaterally removal of excess tissue ventral to the spinal cord along the red line (a) before, and (b) after. (G) Cutting of the ventral spinal cord at the two locations indicated by the red lines. On the rostral side, cutting of the floating ventral spinal cord transversely above C1 where the first GFP-positive anterior horn projects. Cutting of the caudal end of the spinal cord transversely at the upper boundary of the lower limb. Once these cuts are made, the cervical (C1) through lumbar (L2-L3) portion of the ventral spinal cord can be dissected away. (H) Final view of the ventral spinal cord containing GFP-positive SMN columns. Please click here to view a larger version of this figure.
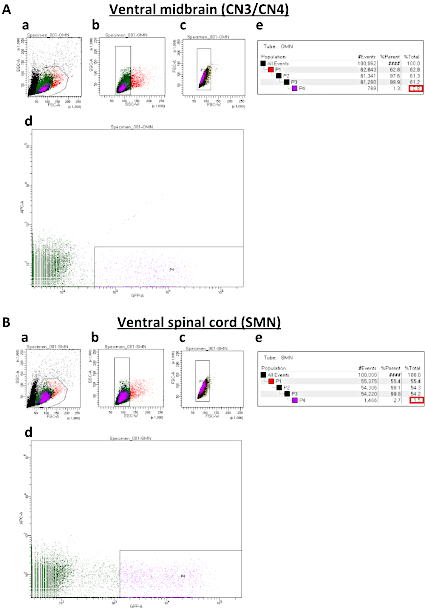
Figure 3: Representative sort plots of ventral midbrains (A) and ventral spinal cords (B). (Aa and Ba) Forward Scatter Area (FSC-A) versus Side Scatter Area (SSC-A) sorted plot before exclusion of debris and dead cells. (Ab, Bb, Ac, Bc) Sorted plots for exclusion of cell clumps (b) and doublets (c) based on Width (SSC-W) versus SSC-A and Forward Scatter Width (FSC-W) versus FSC-A, respectively. (Ad and Bd) Sorted plots to isolate IslMN:GFP -positive motor neurons. In order to obtain a pure culture, the GFP gate must be set higher for SMNs (Bd) than for CN3s/CN4s (Ad). (Ae and Be) Percentages of cells gated for collection by FACS sorting. %Parent represents the percentage of cells in the current gated population relative to the number of cells in the previous gated cell population, whereas %Total represents the percentage of gated cells relative to total cells. Expected percentages of GFP-positive cells as compared to total cells (boxed in red) are 0.5−1.5% for CN3/CN4 and 1.5−2.5% for SMN. If the dissection was performed successfully, these percentages can be used as a benchmark to set up the GFP-positive gate in (Ad and Bd). Please click here to view a larger version of this figure.
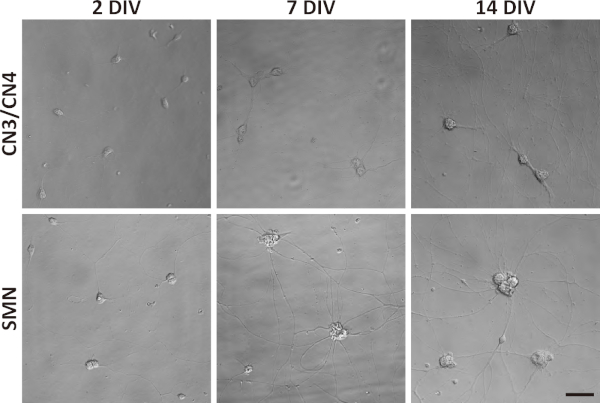
Figure 4: Phase-contrast images of primary CN3/CN4 and SMN monocultures at 2, 7, and 14 DIV. Representative differential interference contrast images of primary CN3/CN4 and SMN cultures were captured at 2, 7, and 14 DIV with inverted fluorescence microscope using corresponding image acquisition and processing software and 40x objectives. Neuronal processes became thicker and longer by 14 DIV. Neuronal cell body sizes became enlarged and tended to aggregate in long-term cultures, especially for SMNs. Both cultures can be maintained at least 14 DIV. Scale bar = 50 µm. Please click here to view a larger version of this figure.
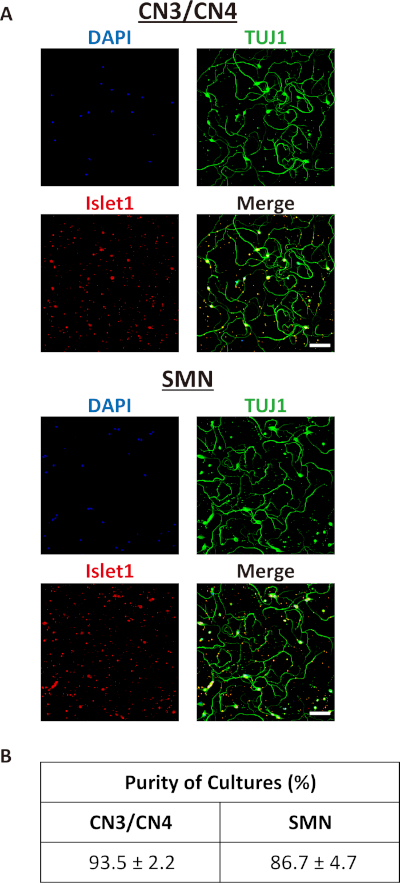
Figure 5: Characterizations of isolated E11.5 mouse CN3/CN4 and SMN cultures. (A) Representative immunocytochemistry images of E11.5 mouse CN3s/CN4s (top) and SMNs (bottom) cultured for 2 DIV. Immunofluorescence labeling with the neuronal marker TUJ1 (green) and the motor neuron marker Islet1 (red) performed to analyze neurons and nuclei were counterstained with DAPI (blue). Almost all the cultured cells were motor neurons (TUJ1+, Islet1+). Images were captured with an inverted fluorescence microscope using corresponding image acquisition and processing software and 20x objectives. Samples were imaged and processed to achieve maximum signal intensity without saturated pixels. All of the microscopic work and image processing in the following figures were performed in these conditions unless otherwise specified. Scale bar = 100 µm. (B) The purities of E11.5 mouse CN3/CN4 and SMN cultures at 2 DIV. The purities of CN3/CN4 and SMN cultures were 93.5 ± 2.2% and 86.7 ± 4.7%, respectively. Dead neuronal cell bodies were assessed by screening for pyknotic nuclear morphology and membrane swelling. Neuronal processes were classified as degenerating processes when signs of beading and swelling were observed. Cells with neither cell body death nor degenerating processes were considered viable non-motor neurons (TUJ1+, Islet1–) or viable motor neurons (TUJ1+, Islet1+)33. The purities of motor neuron cultures were calculated as the number of viable motor neurons divided by the total number of viable non-motor neurons plus viable motor neurons. Values represent the mean ± SEM of three separate experiments. Not significant (p > 0.05) by Student's t test. Cell counting was performed manually under 20x magnification. Abbreviations: SEM = standard error of the mean. Please click here to view a larger version of this figure.
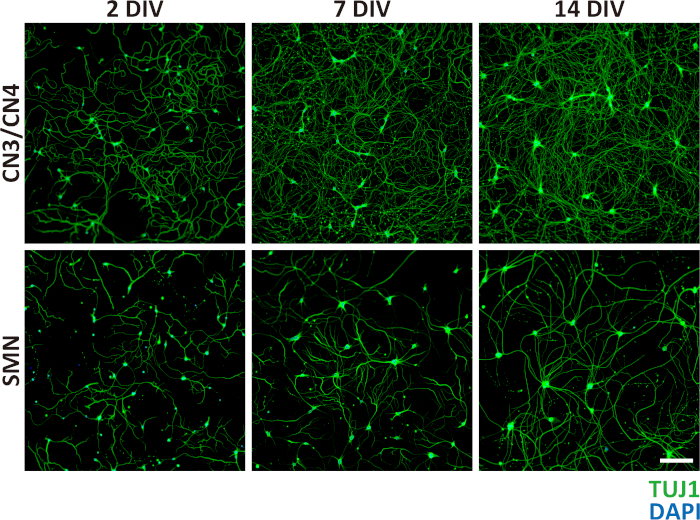
Figure 6: Representative immunocytochemistry of primary CN3/CN4 and SMN monocultures at 2, 7, and 14 DIV. Primary CN3/CN4 and SMN cultures were analyzed at 2, 7, 14 DIV by immunofluorescence labeling with TUJ1 (green), and nuclei were counterstained with DAPI (blue). Neuronal processes become thicker and longer by 14 DIV. Neuronal cell body sizes became enlarged and tended to aggregate in long-term cultures, particularly for SMNs. Both CN3/CN4 and SMN cultures can be maintained at least 14 DIV. Images were captured under 10x magnification. Scale bar = 200 µm. Please click here to view a larger version of this figure.
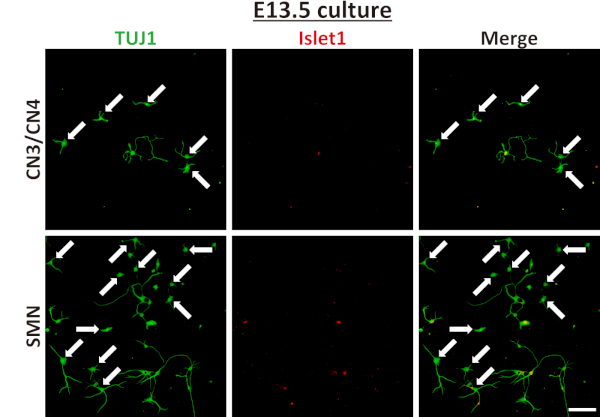
Figure 7: Characterization of E13.5 mouse CN3/CN4 and SMN isolated cultures. E13.5 CN3/CN4 and SMN were isolated and cultured using this protocol and analyzed at 2 DIV by immunofluorescence labeling with TUJ1 (green) and Islet1 (red), and the nuclei were counterstained with DAPI (blue). Many non-motor neuronal cells (TUJ1+, Islet1–) were present (arrows) resulting in a drastic decrease in both CN3/CN4 and SMN purity, with the decrease more pronounced in SMN cultures. Images were captured under 20x magnification. Scale bar = 100 µm. Please click here to view a larger version of this figure.
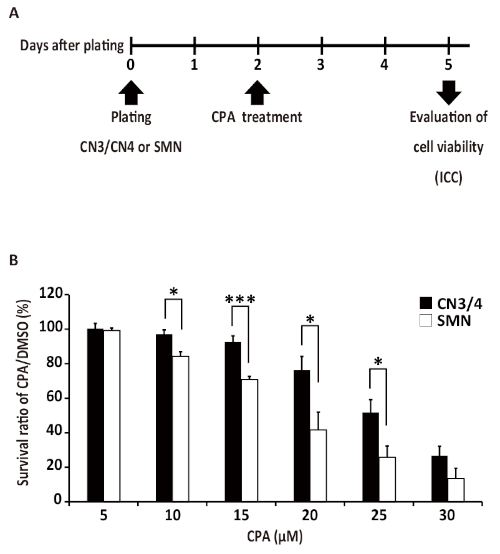
Figure 8: Representative application of primary motor neuron culture demonstrating that CN3s/CN4s are selectively resistant to ER stress induced by CPA. (A) Experimental outline: primary CN3/CN4 and SMN monocultures were treated with CPA or vehicle control (DMSO) at 2 DIV and cell viabilities were evaluated through immunocytochemistry analysis after 3 days of treatment. This outline has been modified from published work31. (B) Quantification of survival ratios of CN3s/CN4s and SMNs treated with 5−30 µM CPA for 3 days from 2 DIV. Neurons were analyzed by immunofluorescent labeling of cells with TUJ1, and nuclei were counterstained with DAPI. Survival ratios were calculated as the number of viable cells (see Figure 5B legend) in drug-treated wells divided by the number of viable cells in wells containing vehicle alone (DMSO). Cell counting was performed manually under 20x magnification. Values represent the mean ± SEM of four separate experiments. *p < 0.05; ***p < 0.005 by Student's t test. This figure has been modified from previously published work31. Please click here to view a larger version of this figure.
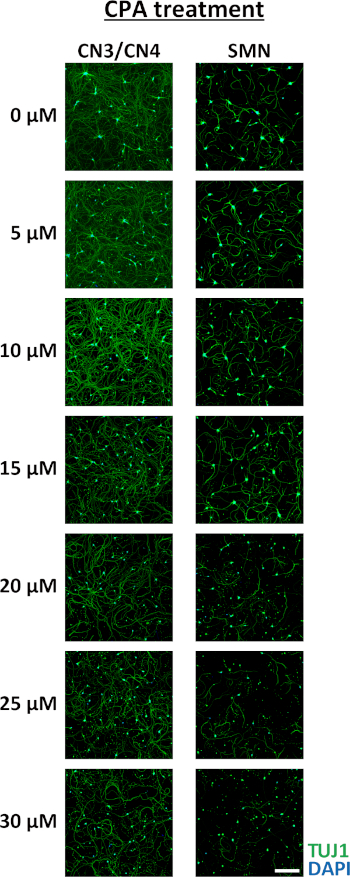
Figure 9: Representative immunocytochemistry of primary CN3/CN4 and SMN monocultures after a 3 day exposure to increasing concentrations of CPA beginning at 2 DIV. Neurons were analyzed by immunofluorescent labeling of cells with TUJ1 (green) and nuclei were counterstained with DAPI (blue). Primary CN3s/CN4s were more resistant to CPA treatment than primary SMNs. Images were captured under 10x magnification. Scale bar = 200 µm. Please click here to view a larger version of this figure.
| Number of midbrains (X) | Papain | Albumin-ovomucoid | Hibernate E | |||||||||
| 10 ≤ X ≤20 | 200 μL | 100 μL | 600 μL | |||||||||
| 20 < X ≤30 | 300 μL | 150 μL | 700 μL | |||||||||
| 30 < X ≤40 | 400 μL | 200 μL | 800 μL | |||||||||
| Number of spinal cords (Y) | Papain | Albumin-ovomucoid | Hibernate E | |||||||||
| 3 ≤ Y ≤5 | 200 μL | 100 μL | 500 μL | |||||||||
| 5 < Y ≤10 | 400 μL | 200 μL | 800 μL | |||||||||
| 10 < Y ≤15 | 600 μL | 300 μL | 1200 μL | |||||||||
Table 1: Appropriate volumes of papain, albumin-ovomucoid, and final suspension used in dissociation steps. The appropriate volumes of papain and albumin-ovomucoid to be used with various numbers of ventral midbrain and ventral spinal cord tissues were modified from the manufacturer's instructions after several rounds of optimization. Because tissues are subject to stress during dissociation and sorting, a pooled collection of more than 10 ventral midbrains and more than three ventral spinal cords is recommended. The volume of papain was determined by considering the balance between effective dissociation and the stress of this procedure. The volume of albumin-ovomucoid inhibitor solution is half of that of papain. The appropriate volume of Hibernate E final suspension was determined such that cell density does not exceed 107 cells/mL, but the cells do not become excessively diluted.
