Um grande número de eventos de motilidade dentro das células eucarióticas são mediados pela função das proteínas motoras moleculares. Esses motores se movem ao longo dos filamentos citoesqueléticos, filamentos de actin e microtúbulos (MTs), e convertem a energia química da hidrólise ATP em forças cinéticas e mecânicas necessárias para conduzir a motilidade biológica dentro das células. O S. cerevisiae Cin8, baseado em MT, é uma proteína motora bipolar, homotetrameric quinase-5 que cruza e desliza MTs de eixo1. O Cin8 executa funções essenciais durante a mitose, na montagem do eixo 2,3,4 e no alongamento do eixo durante a anafase 5,6,7. Anteriormente, havia sido demonstrado que o Cin8 é um motor bidirecional, que muda a direcionalidade em diferentes condições experimentais. Por exemplo, sob altas condições de resistência iônica, motores Cin8 únicos movem-se em direção à extremidade inferior dos MTs, enquanto em clusters, em ensaios de deslizamento MT multimotores, e entre MTs antiparalhais, os motores Cin8 movem-se principalmente em direção às extremidades mais altas dos MTs 8,9,10,11,12 . Esses achados foram altamente inesperados por várias razões. Primeiro, o Cin8 carrega seu domínio motor catalítico no amino-terminus e tais motores foram previamente acreditados como exclusivamente mais-end dirigidos, enquanto Cin8 foi mostrado ser menos-fim direcionado ao nível de molécula única. Em segundo lugar, acreditava-se que os motores de cinesina eram unidirecionais, seja de ponta ou de ponta, enquanto o Cin8 mostrou-se bi-direcional, dependendo das condições experimentais. Finalmente, por causa da orientação em MT no fuso mitótico, o papel clássico dos motores cinesina-5 na separação de postes de fuso durante a montagem do fuso e da anfífase B só poderia ser explicado por sua motilidade direcionada plus-end nos MTs que eles cruzam 1,13. Após os primeiros relatos sobre a bidirecionalidade do Cin8, alguns outros motores de cinesina foram demonstrados como bi-direcionais 14,15,16, indicando que a motilidade bidirecional dos motores de cinesina pode ser mais comum do que se acreditava anteriormente.
Foi relatado anteriormente que nas células, o Cin8 também se move de forma bidirecional8, apoiando a noção de que a motilidade bidirecional de alguns motores cinesin-5 é importante para suas funções intracelulares. Além disso, uma vez que os três motores cinesin-5 que foram relatados como bidirecionais são de células fúngicas, um possível papel para a bidalidade bi-direcionalidade dos motores cinesin-5 foi recentemente proposto nessas células10. De acordo com este modelo, em mitose fechada de células fúngicas, onde o envelope nuclear não quebra durante a mitose, os motores de quinase-5 fornecem a força inicial que separa os polos do eixo antes da montagem do eixo. Para realizar esta tarefa, antes da separação do polo de fuso, os motores kinesin-5 localizam-se perto dos polos de fuso, por sua motilidade direcionada sem fim em MTs nucleares únicos. Uma vez nesta posição, os motores kinesin-5 agrupam, alternam direcionalidade, capturam e cruzam MTs de postes de fuso vizinhos. Posteriormente, os motores kinesin-5 fornecem a separação inicial dos polos por motilidade dirigida plus-end nos MTs que eles cruzam. Por este modelo, tanto a motilidade dirigida por fim em MTs único quanto a motilidade direcionada plus-end em MTs transversais durante o deslizamento antiparal são necessárias para que os motores fúngicos kinesin-5 realizem seus papéis na montagemdo eixo 1,13.
O objetivo geral do método descrito é obter kinesina-5 Cin8 marcada por FFP de alta pureza e realizar ensaios de motilidade de molécula única (Figura 1) enquanto analisa separadamente a motilidade de moléculas únicas e aglomerados de Cin8. A separação entre moléculas únicas e aglomerados é importante, uma vez que um dos fatores que foram demonstrados para afetar a direcionalidade do Cin8 é o seu acúmulo em clusters nos MTs10,12. Ensaios alternativos de motilidade, como o deslizamento de superfície mt e ensaios deslizantes de MT não fornecem informações sobre a atividade de proteínas motoras únicas 17,18. Os robustos métodos de ensaio e análise de motilidade de molécula única descritos aqui foram aplicados com sucesso para caracterizar diferentes aspectos dos motores kinesin-5, Cin8 e Kip1 10,11,12,14,19,20.
Aqui, um protocolo detalhado é apresentado para a superexpressão e purificação do Cin8, polimerização de MTs e o ensaio de motilidade de molécula única. Além disso, também são descritas as análises para diferenciar moléculas e aglomerados únicos de Cin8 e determinar velocidades motoras e cluster únicas por deslocamento médio (MD) e análise média de deslocamento quadrado (MSD). Este protocolo tem como objetivo ajudar os pesquisadores a visualizar todas as etapas dos procedimentos e auxiliar na solução de problemas desse tipo de ensaio.
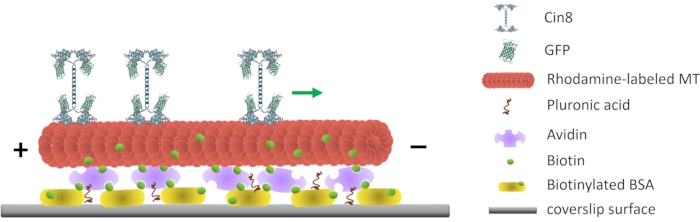
Figura 1: Representação esquemática do ensaio de motilidade de molécula única. Os MTs fluorescentes biotinilados são anexados à superfície de vidro, revestidos com o Avidin que interage com o biotinilado-BSA conectado à superfície. O arqueiro verde representa a direção de movimento de moléculas cin8 únicas sob altas condições de resistência iônica. +/- representar a polaridade do MT. Clique aqui para ver uma versão maior desta figura.
