Resonance Energy Transfer (RET) is a non-radiative interaction between a luminescent donor and a fluorescent acceptor, whereby the energy from the exited donor is capable of inducing a fluorescence response from the acceptor through a strong dipole-dipole interaction13. RET, which has been described using fluorescent23, chemiluminescent24, and bioluminescent13 donors, principally requires: 1. Strong spectral overlap between the donor emission and accepter excitation spectra; 2. Appropriate rotational alignment between the two entities; and 3. A working distance no greater than 0.5- to 2-times the Förster radius, R0, between the donor and acceptor17. Contrasting with RET, FUEL occurs when a luminescent source, such as a bioluminescent bacteria, emits a photon that is absorbed and re-emitted by a second entity, such as a fluorophore or a fluorescent nanoparticle, red-shifting the emission spectrum of the original luminescent source. Thus, FUEL follows a standard excitation-emission process akin to standard epifluorescent conditions, yet without the use of a focused excitation. Evidence of this can be readily observed by the simple mixing of luminescent bacteria and highly-fluorescent quantum dots. In the presence of the Vibrio sp, a significant increase in red signal is observed when QD705 were also in solution compared to a non-fluorescent polystyrene microsphere control (Figure 1A). The components necessary to create bioluminescence, essentially the aldehyde substrate, luciferase, and ATP, are all produced and contained within the cytoplasm. Furthermore, the enzymatic luminophore production occurs within the bacterial cytoplasm (Figure 1B). As has been reported elsewhere, the distance between the inner and outer membrane of bacteria is typically greater than 30 nm25-27, a distance that does not allow for significant RET to occur. As well, the fluorescent nanoparticles do not cross into the bacterial cytoplasm since there is no endocytosis or other means of uptake. Together, these constraints indicate that FUEL is the dominant excitation-emission phenomenon that occurs when luminescent bacteria are mixed with fluorescent nanoparticles.
As was previously shown, FUEL exists beyond the range of RET. To investigate the FUEL dependence on distance, standard reduced volume spectrophotometric cuvettes can be used, with one containing a fluorophore solution, a second containing an appropriate control that can control for scatter, and the third containing an aliquot of fresh luminescent solution. It is essential that the central cuvette be enveloped using black tape or another opaque material, save for at least two identical optical windows located on opposite faces, to reduce any potential light contamination from the luminescent source. Further, the two remaining cuvettes need to be placed equidistantly onto either side of the central cuvette. Finally, a fresh luminescent solution, using bacteria or chemiluminescence, should be used with each experiment to ensure maximum light production. Upon appropriate placement, acquire the luminescence signal under the desired filters. The Total Light and 710-730 nm filters were used in this case, though only the data from the latter is shown. After each acquisition, increase incrementally the face-to-face distance from 1.0 cm to 3 cm (Figure 2). Finally, normalize all the data to the brightest point. In the examples used here, this occurred at the shortest distance (D = 1 cm) between the luminescent source and the cuvette containing the QD705. Using this approach, viable FUEL signal can be observed up to the final acquisition suggesting that, in the absence of any optical absorber, FUEL can occur at distances beyond any possible resonance energy transfer and can only be explained to be a purely radiative effect. Fitting the data reveals a decrease in signal as a function of distance D following a D-1 to D-2 dependence. Depending upon the geometric configuration, the former corresponds to the capacitor distance dependence and the latter to the inverse square law for point sources. This result is therefore consistent with our overall acquisition setup, given the size of the cuvette aperture and the distance between the luminescent source and the fluorophore.
FUEL not only applies to quantum dots, but also can be observed using a wide range of fluorophores ranging from the Alexa series to fluorescent microspheres (Table 2). In order to be comparable to the control, equal concentrations of the various fluorophores should be used and the total surface area of the nanoparticles held constant (Table 1). By comparing the fluorophores and nanoparticles to their appropriate controls (PS or PS with non-fluorescent polystyrene microspheres), a significant increase in signal is observed at the emission maximum of each fluorescent entity. The largest relative increase in signal was found to occur where the fluorescent emission maximum was farthest from the luminescent emission maximum. This is most likely due to the increased specificity of the fluorescent signal compared to the broad emission spectrum of the luminescent source. On the contrary, the least reliable FUEL signal was found when the two emission maxima were not well separated. Interestingly, a discernable FUEL signal was found with the Yellow microspheres even though the spectral difference is minimal, due most likely to the substantial quantity of the fluorophore present per bead (350 fluorescein equivalents). The results shown here indicate that under appropriate conditions FUEL can be achieved with a variety of fluorophores, which enables the tailoring of probes chosen for more relevant applications under both in vitro and in vivo conditions. The significance of the observed FUEL signal was determined using a standard two-tailed Student's t-test. As such, only the Alexa555 was found to be incompatible with the luminescent source used.
In order to explore the effects of targeting on FUEL, biotinylated antibodies were used to target luminescent bacteria and streptavidin-linked QDs. It is important that the bacteria or luminescence source provide a layer thick enough to minimize the possibility of RET between the luminophore and the corresponding fluorophore. After incubation and washing to remove unbound antibodies, the biotin-labeled bacteria are then exposed to either streptavidin-conjugated QD705 or non-functionalized QD705 as the control, the solutions divided and one set exposed to further washes in order to remove any non-adhered QD705. Here, for all four conditions, the resulting luminescence was observed under the Total Light, 490-510 nm, and 710-730 nm filters, though only the latter two filters are used for the data analysis. The presence of the QD705 was compared using 450-480 nm excitation and 710-730 nm emission filters. Upon investigation we have found little to no difference in red-shifted signal when the solutions were left in an unwashed state (Figure 3). The resulting fluorescence signal under this condition is also found to be quite similar, suggesting that an equal number of QD705 was present under both the targeted and non-targeted state. However, washing the samples to remove any unbound QD705 provides a nearly two-fold increase in relative red signal for the targeted bacteria compared to their non-targeted control. Investigation by fluorescence can be used to verify the presence or absence of the QD705. Comparatively, the targeted washed condition resulted in a fluorescence intensity that was almost three times less than the unwashed condition, yet the resulting red-shift decreased by only 30%, suggesting that under purely bound conditions the targeting of the bacteria will result in an increase in red-shifted emission. This strongly implicates the utility of targeted FUEL for future applications.
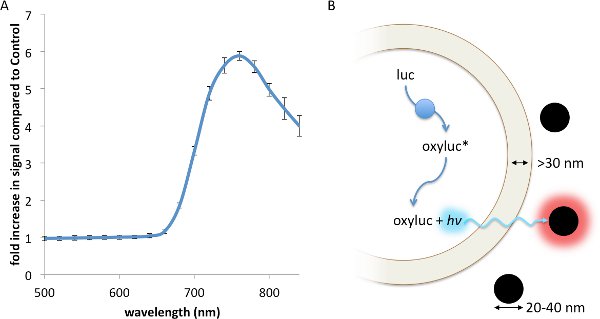
Figure 1. A FUEL interaction between luminescent bacteria and commercially available quantum dots leads to an increase in red photon production. Two spectrophotometric cuvettes were filled with solutions containing 100 μl aliquots of fresh V. fischeri culture with an OD600 of 1-1.5, 895 μl of PS, and either 5 μl of QD705 or physiological saline (PS), before being placed into an IVIS Spectrum and the emission spectrum acquired. A significant increase in red signal is achieved due to the presence of the QD705, as can be noted by the increase in detected photons•sec-1•cm-2 (p•sec-1•cm-2) compared to the control under the 710-730 nm emission filter (A). Here, the dual membrane of the bacteria excludes the interaction of the luminescent moieties (blue circles) and the QD705 (black circles). The former are found only in the bacterial cytoplasm while the latter are freely distributed in the bulk solution (B). N=3 for each bacterial solution. Please click here to view a larger version of this figure.
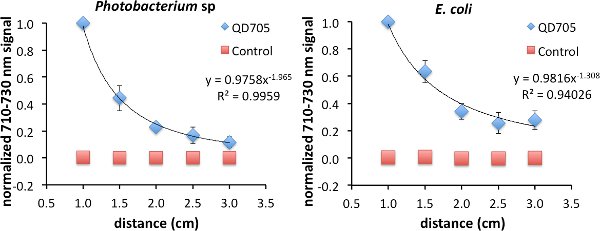
Figure 2. Evidence of FUEL occurring over distances completely excluding resonance energy transfer. Spectrophotometric cuvettes were filled with solutions containing either 50 μl of QD705 and 950 μl of physiological saline (PS) or 97.2 μl of non-fluorescent 48 nm polystyrene microspheres and 902.8 μl of PS, and placed equidistantly on opposite sides of a central black cuvette containing 1 ml of fresh luminescent Photobacterium sp at an OD600 of 1-1.5. The central cuvette had two identical optical windows allowing the emitted photons to freely disperse. Acquiring images at distances (D) ranging from 1.0 cm to 3 cm, the observed production of red light decreased as a function of distance displaying evidence that FUEL can occur at distances not achievable by resonance energy transfer. The intensity appeared to have D-2 dependence, similar to the inverse square law (left). The same protocol was followed for E. coli (right). The resulting trend lines hold form to the concept that the bacteria are acting as point sources of light. A total of three independent distance measurements were acquired with each using a unique subculture. Error bars are present at each point. In some cases the error bars were smaller than the symbols used that indicate the normalized intensity. Please click here to view a larger version of this figure.
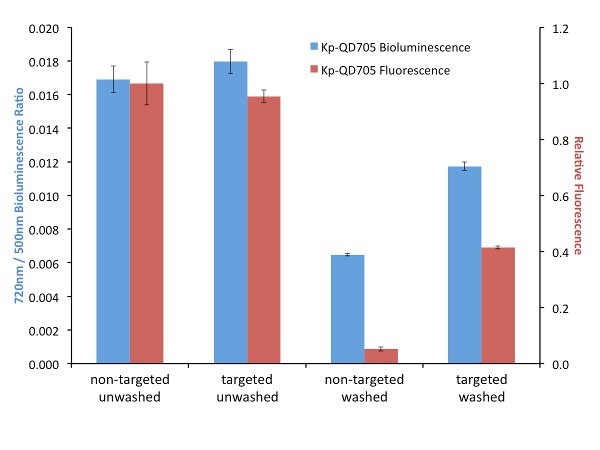
Figure 3. A comparison between targeted (specific) and non-targeted (bulk solution) FUEL. K. pneumoniae were functionalized with biotinylated antibodies and then exposed to either streptavidin-labeled (Targeted) or non-labeled (Non-Targeted) QD705. The resulting solutions were equally divided and one set washed three times with PBS before being resuspended into its initial volume in PBS. The solutions were then dispensed into individual wells of a black 96-well plate and the bioluminescence observed under the 490-510 nm (500 nm) and 710-730 nm (720 nm) emission filters (indicated as blue bars). The p•sec-1•cm-2 found from the 720 nm filter for each well was normalized by the respective p•sec-1•cm-2 bioluminescence intensity from the 500 nm of the same well. The relative fluorescence intensity resulting from an epifluorescent excitation was determined using 450-480 nm excitation and 710-730 nm emission filters (indicated as red bars). Little to no difference in bioluminescent or fluorescent signal was observed under the unwashed conditions (Non-Targeted Unwashed and Targeted Unwashed). This suggests that the bound and free-floating QD705 were equally excited by the bioluminescent photons. Upon washing, nearly a two-fold difference in relative red signal was observed for the QD705-labeled bacteria (Targeted Washed) compared to the control (Non-Targeted Washed). The absence of QD705 in the Non-Targeted Washed was confirmed by the lack of fluorescent signal and verified the QD705 labeling in the Targeted Washed state. The data is from four independent cultures of K. pneumoniae. For clarity, the legend indicates the source of the QD705 excitation. Please click here to view a larger version of this figure.
| Fluorophore | Concentration | μl | PS (μl) | λmax ex/em (nm) | Emission Filter (nm) |
| Alexa 555 | 37.2 µM | 4.77 | 995.23 | 555/565 | 570 – 590 |
| Alexa 568 | 37.2 µM | 4.77 | 995.23 | 578/603 | 590 – 610 |
| Alexa 633 | 37.2 µM | 4.77 | 995.23 | 632/637 | 650 – 670 |
| Alexa 700 | 37.2 µM | 4.77 | 995.23 | 702/723 | 710 – 730 |
| Nonfluorescent | 2.62% solids | 9.72 | 990.28 | ||
| µSph Pink | 5% solids | 4.77 | 995.23 | 580/605 | 610 – 630 |
| µSph Yellow | 5% solids | 4.77 | 995.23 | 505/515 | 510 – 530 |
| QD705 | 2 µM | 5 | 995 | 465/705 | 710 – 730 |
| QD800 | 2 µM | 5 | 995 | 465/705 | 790 – 810 |
Table 1. Properties of fluorophores used throughout the FUEL demonstrations.
| Control | filter | fluorophore | Control | p value | |||
| p·sec-1·cm-2 | SD | p·sec-1·cm-2 | SD | ||||
| A555 | PS | 580 | 1.08 x 107 | 3.31 x 106 | 9.09 x 106 | 1.75 x 106 | 0.233 |
| A568 | PS | 600 | 8.47 x 106 | 4.23 x 106 | 4.49 x 106 | 9.71 x 105 | 0.094 |
| A633 | PS | 660 | 2.40 x 106 | 1.25 x 106 | 7.85 x 105 | 2.39 x 105 | 0.046 |
| A700 | PS | 720 | 5.53 x 105 | 2.46 x 105 | 1.54 x 105 | 6.05 x 104 | 0.026 |
| MSph Yellow | non-fluorescent µspheres | 520 | 1.19 x 108 | 4.85 x 107 | 5.79 x 107 | 1.99 x 107 | 0.057 |
| MSph Pink | non-fluorescent µspheres | 620 | 2.37 x 107 | 1.36 x 107 | 2.16 x 106 | 8.00 x 105 | 0.026 |
| QD705 | non-fluorescent µspheres | 720 | 1.76 x 107 | 7.33 x 106 | 2.08 x 105 | 7.16 x 104 | 0.007 |
| QD800 | non-fluorescent µspheres | 800 | 7.79 x 106 | 4.72 x 106 | 3.60 x 104 | 1.52 x 104 | 0.023 |
Table 2. Identification of fluorophores and fluorescent nanoparticles compatible with FUEL.
