x`Mosquito dissections
Following the protocol, we collected and isolated the ovaries, midgut, salivary glands, and the carcass from two Culex pipiens molestus specimens (including a gravid female) from a laboratory colony. We confirmed clean dissections following the observation of entire (that is, unbroken) and well-isolated organs with no remaining debris under the binocular. The whole body, head, and thorax, dissected salivary glands, ovaries, and midgut of a Culex pipiens molestus specimen are shown in Figure 2. As expected, the midgut and salivary glands were an order of magnitude smaller than the mosquito ovaries. The eggs of a gravid Culex specimen, together with its midgut and Malpighian tubules, are presented in Figure 3. Of note, dissections are more prone to fail when the material is not fresh and tissue is likely to break (Figure 4B,C). We therefore suggest dissecting material right after collection (within less than 12 h) when possible as tissues are still elastic (Figure 4A). Similarly, dissection of frozen material can be performed but there is a much higher risk of failure and cross-contamination between organs due to fragile tissues (Figure 4).
Microbiome data
In addition, we collected and separated the midguts and ovaries from three individual Culex quinquefasciatus specimens from Noumea, New Caledonia, following the same procedure. We extracted DNA from each organ, prepared samples for whole genome sequencing, and performed microbiome data analyses as detailed in Supplemental File 1. A bacterial taxonomic diversity analysis on unassembled quality-filtered short reads using PhyloFlash12 showed distinct dominant taxa in midguts compared to ovaries (Figure 5A). Notably, ovarian bacterial communities were dominated by Wolbachia, with the additional presence of a Spirochaetaceae phylotype in the ovaries of individual 3, while midgut communities exhibited a wider diversity, including Gammaproteobacteria, Spirochaetaceae, and Firmicutes. From the same sequencing data, we reconstructed four Metagenome-Assembled Genomes (MAGs) with completion > 80% and redundancy < 5%, belonging to classes Spirochaetia, Alpha-, and Gammaproteobacteria (Table 1).
As expected, MAGs reconstructed herein did not cover the full taxonomic diversity predicted by the PhyloFlash results due to the specific shortcomings of genome reconstruction using metagenomic short-reads. The MAG assigned to Wolbachia (Ref. MAG 3 in Table 1) was detected in all ovaries and two of the midguts and had higher coverage in the ovaries (Figure 5B). We also reconstructed two MAGs belonging to the Enterobacteriaceae family, including genus Pantoea (Ref. MAG 1 and 2 in Table 1), in the midguts obtained from Culex individuals 2 and 3, that were not detected in the corresponding ovaries (Figure 5B). Finally, we also reconstructed one Spirochaetaceae bacterial genome, Ref. MAG 4 (Table 1), assigned to genus BR149 that was successfully isolated from Culex pipiens midguts by Graña-Miraglia and colleagues13. Interestingly, this MAG was detected in the midguts of individuals 1 and 3, as well as in the ovaries of individual 3 (Figure 5B).
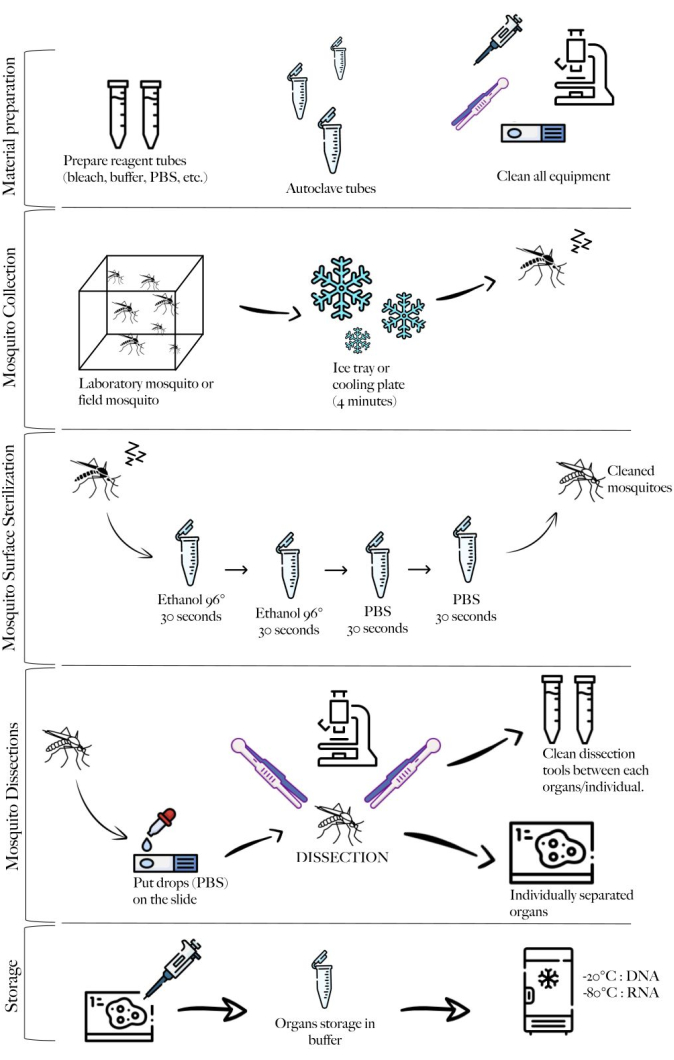
Figure 1: Schematic of the method summarizing the different steps. Material preparation, mosquito collection, mosquito cleaning, mosquito dissections, and storage. Please click here to view a larger version of this figure.
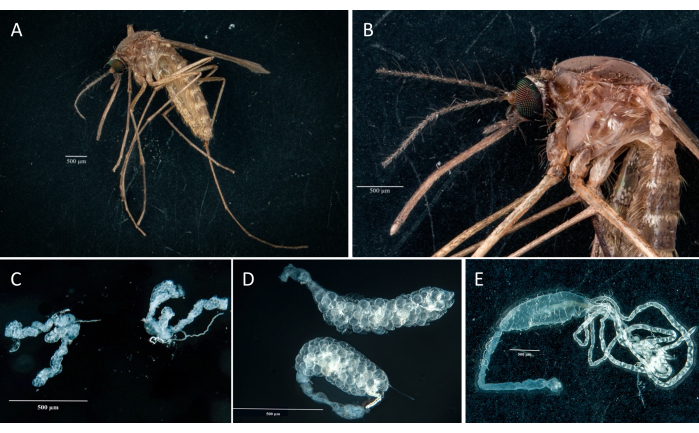
Figure 2: Culex pipiens molestus female. (A) Whole body. (B) Head and thorax. (C) Dissected salivary glands. (D) Dissected ovaries. (E) Dissected gut with Malpighi tubules. Scale bars = 500 µm. Please click here to view a larger version of this figure.
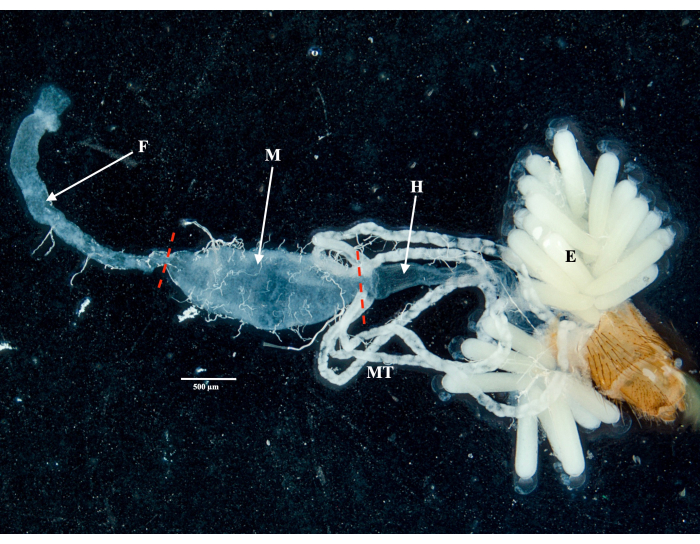
Figure 3: Dissected abdomen of a gravid Culex pipiens molestus female. The foregut, midgut, hindgut, Malpighian tubules, and eggs of the specimen are shown. Red dashes indicate where to cut to separate the midgut from the foregut and hindgut. Scale bar = 500 µm. Abbreviations: F = foregut; M = midgut; H = hindgut; E = eggs; MT = Malpighian tubules. Please click here to view a larger version of this figure.
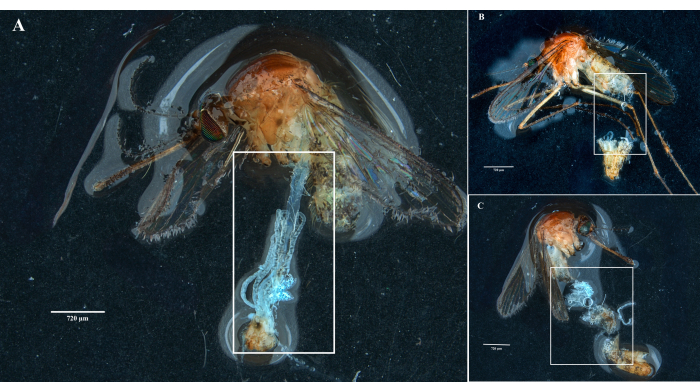
Figure 4: Dissection of fresh and frozen Culex mosquito specimens. White rectangles illustrate (A) mosquito tissues with intact organs from a freshly dissected specimen and (B,C) broken biological material from specimens that have been frozen before dissection. Scale bars = 720 µm. Please click here to view a larger version of this figure.
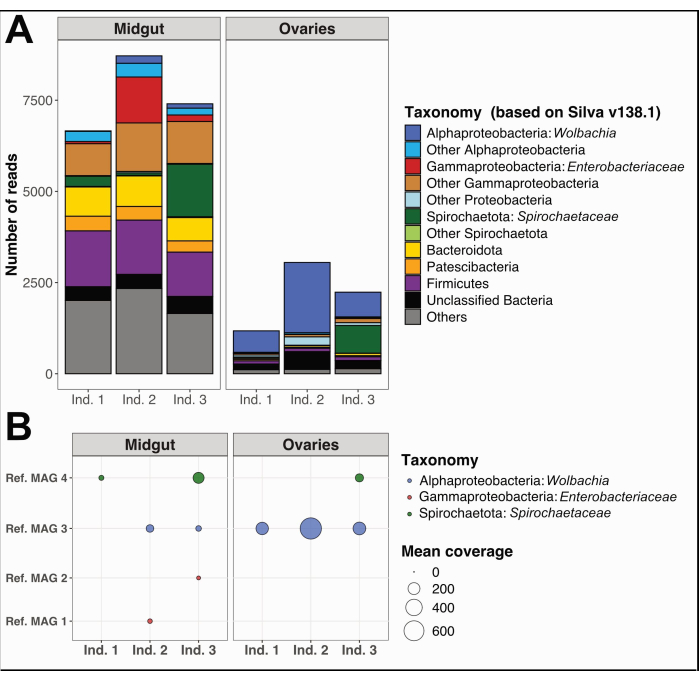
Figure 5: Example microbiome analysis on three Culex quinquefasciatus individuals. (A) Visualization of bacterial diversity estimated through the extraction of SSU rRNA reads with PhyloFlash12 in midguts and ovaries of the three specimens. (B) Mean coverage of the reconstructed MAGs over the samples. Abbreviations: MAG = Metagenome-Assembled Genome; Ind = Individual. Please click here to view a larger version of this figure.
| MAG | Ref. MAG 4 | Ref. MAG 2 | Ref. MAG 1 | Ref. MAG 3 |
| Length (bp) | 1,287,790 | 4,910,866 | 4,751,276 | 1,298,266 |
| Number of contigs | 9 | 162 | 149 | 123 |
| GC % | 34.05 | 55.45 | 54.05 | 34.15 |
| Completion (%) | 84.5 | 97.18 | 98.59 | 91.55 |
| Redundancy (%) | 0 | 2.82 | 4.22 | 0 |
| Domain | Bacteria | Bacteria | Bacteria | Bacteria |
| Phylum | Spirochaetota | Proteobacteria | Proteobacteria | Proteobacteria |
| Class | Spirochaetia | Gammaproteobacteria | Gammaproteobacteria | Alphaproteobacteria |
| Order | WRBN01 | Enterobacterales | Enterobacterales | Rickettsiales |
| Family | WRBN01 | Enterobacteriaceae | Enterobacteriaceae | Anaplasmataceae |
| Genus | BR149 | Pantoea | – | Wolbachia |
Table 1: Reconstructed MAGs from the three Culex quinquefasciatus individuals. Genome size, number of contigs, proportion of GC, estimates of completion, and redundancy based on the single-copy core gene collection available in Anvi'o17 and taxonomy obtained using GTDB18.
Supplemental File 1: Detailed example procedure for microbiome data analysis starting from sample collection, DNA extraction, and whole genome sequencing followed by the bioinformatic workflow for genome reconstruction and estimation of prokaryotic and eukaryotic read proportion Please click here to download this File.
