Quantitative methods are described to study protein-ligand interactions of vitamin A membrane receptors and photoreceptor opsin with their respective physiological ligands. The recombinant mouse RBP4 should be expressed in E. coli and the purified protein used as a conjugated ligand on a SPR Chip. The chemically synthesized RBPR2, STRA6, and mutant S294A RBPR2 "SYL motif RBP4 interacting extracellular site" of a ~40 amino acid peptide is used as analyte at various concentrations to measure the kinetics of binding and equilibrium saturation Kd of interaction (Figure 1 and Figure 2).
Recombinant RBP4 protein, as a ligand for RBPR2 and STRA6, showed a binding affinity towards the interacting sites in these peptides, measured at Kd ~22.38 µM and ~26.73 µM, respectively. The amino acid motif "SYL" plays an essential role in stabilizing these interactions both in RBPR2 and STRA6, and as predicted, the mutant peptide S294A had increased Kd of ~114.9 µM, suggesting an increased copy number of mutant-RBPR2 peptide requirements for reaching binding saturation (Figure 3). Retinol uptake in the eye via STRA6 and conversion into various retinoids to the final conversion of 11-cis retinal as chromophore with binding to Lysine 296 residue of Rhodopsin makes a functional receptor for photon sensing and vision. The 11-cis retinal covalent binding to protein Opsin to form Rhodopsin exhibits certain characteristic absorbance features. Protein absorbance occurs at 280 nm, and free Retinol absorbance is at ~325 nm. However, the holo-Opsin with 11-cis Retinal or Rhodopsin absorbance occurs at 500 nm wavelength.
Any defect in this complex formation because of the low systemic retinol or defect in retinol uptake by a mutation in RBPR2 or STRA6 leads to the accumulation of Free Opsin or apoprotein opsin devoid of chromophore 11-cis retinal, changing the characteristic absorbance shifts to low absorbance at 500 nm. By quantifying the absorbance and using the ratio of absorbance 280 nm and 500 nm wavelength, it is possible to estimate the liganded Opsins or Rhodopsin. Through a brief exposure of light, it is possible to cross-check the photo-isomerization of 11-cis retinal bound Opsin absorbance at 500 nm to all-trans-retinal-bound Opsin (Meta II rhodopsin) absorbance at 380 nm (Figure 4, Figure 5, and Figure 6). In an experimental setup, the chance of artifacts and contaminants is high; the two major sources of artifacts found are unintentional light exposure from the instruments and low-concentration salt wash with failure to remove contaminant molecules from the RPE or lens (Figure 7).

Figure 1: Surface Plasmon Resonance workspace. Materials needed and overview of the SPR instrumentation. (A) SPR Chip loading dock. (A') CM5 Chip with carboxymethylated dextran covalently attached gold surface for immobilization via -NH2, -SH, -CHO, -OH, or -COOH groups. (B) Sample loading robotic injector rack. (B') type 3 rubber caps. (B'') 7 mm plastic vials for sample aliquots. (C,D) Equilibration/running buffer reservoir and waste collection tank. (E) Biacore software control panel. Please click here to view a larger version of this figure.
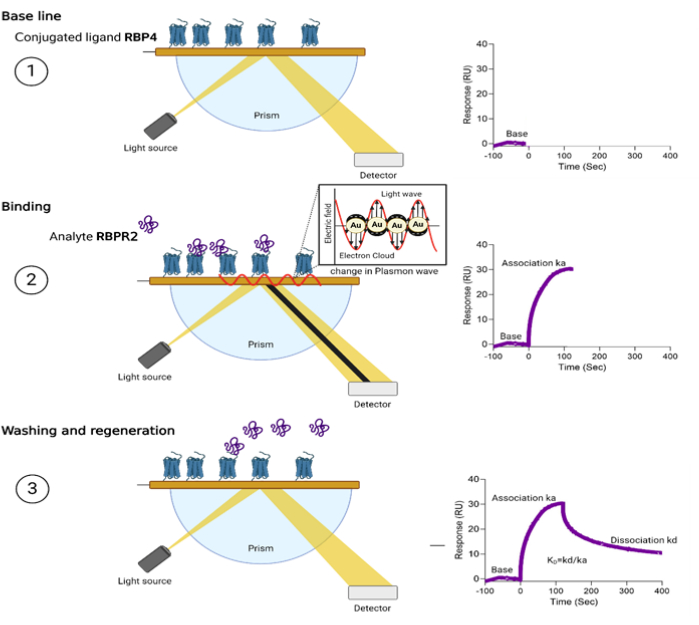
Figure 2: Schematic overview of Surface Plasmon Resonance and the steps of sensorgram acquisitions. Step 1: The CM5 Chip surface immobilized with RBP4 protein with a target of 1,200 response units. Step 2: Various concentrations of RBPR2 peptides are run as analytes, and this biomolecular interaction changes the electron charge density on the gold Chip surface, leading to the change/reduction in light reflection at a specific angle or a resonance angle. Step 3: The washing reverses the surface plasmon phenomenon, leading to the reversal of reflection intensity and the drop of the sensorgram to the baseline. The kinetics of these interactions were further used to determine the association dissociation constants and strength of interactions. Please click here to view a larger version of this figure.
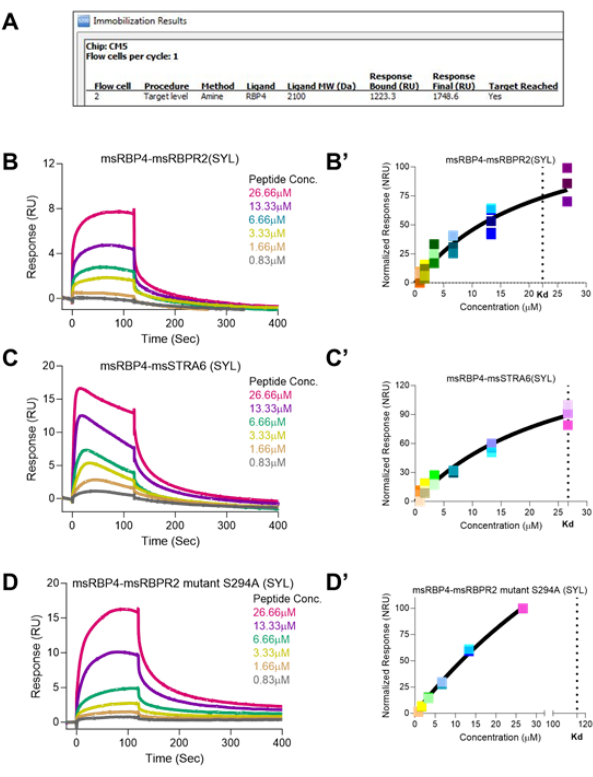
Figure 3: Surface Plasmon Resonance binding studies of individual vitamin A receptor binding domain peptides with its physiological ligand RBP4. (A) The software prompt showing the achieved immobilization of RBP4 on the CM5 Chip surface. (B,C,D) Sensograms of peptide-ligand RBPR2-RBP4, STRA6-RBP4, and mutant S294A RBPR2-RBP4 interactions at various concentrations. (B',C',D') Binding Kinetics and equilibrium Kd determination showing the difference and closeness of binding pattern. Please click here to view a larger version of this figure.
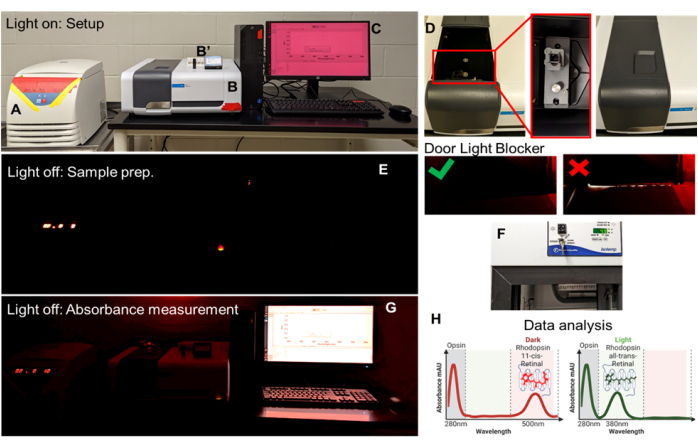
Figure 4: Opsin-11-cis retinal absorbance workspace and overview of the instrumentation. (A) Refrigerated benchtop centrifuge, all LED indicators covered with red tape; (B) UV-visible spectrophotometer, one LED indicator covered with red tape; (B') Rectangular ultra-micro cell, quartz cuvette, and magnetic stand. (C) Scan application software; (D) spectrophotometer compartment cover slide open, cuvette holder for analysis, and compartment cover closed. (E) Light OFF setup with monitor OFF for sample preparation under a single red-light source. (F) Refrigerated rotator for sample preparation and solubilization. (G) Light OFF setup with monitor ON for sample absorbance measurement. (H) Analysis of absorbance in XY-Plot. Please click here to view a larger version of this figure.
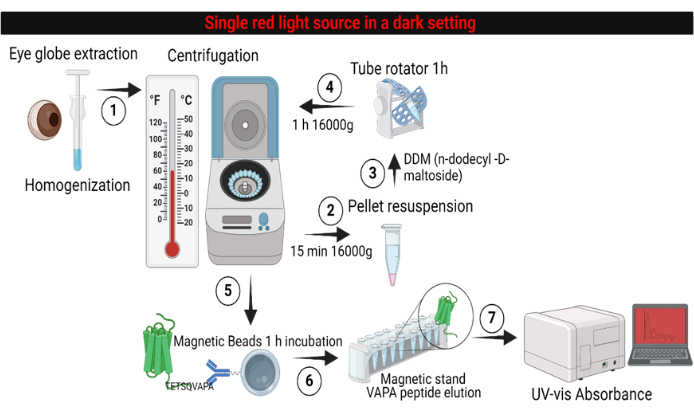
Figure 5: Workflow overview of the Opsin-11-cis retinal absorbance methodology. Step 1: Retinas from dark-adapted mice under a single red-light source are isolated, homogenized, and solubilized to obtain retinal membrane fractions. Steps 2-4: In a sequence of centrifugations, the pellet is solubilized in the absence of detergent and the presence of DDM detergent buffer. Steps 5-6: The solubilized pellet containing retinal photoreceptors and other membrane proteins ready for antibody pulldown and purification. Step 7: The eluted Opsin using VAPA 1D4-peptide ready for absorbance analysis. Please click here to view a larger version of this figure.
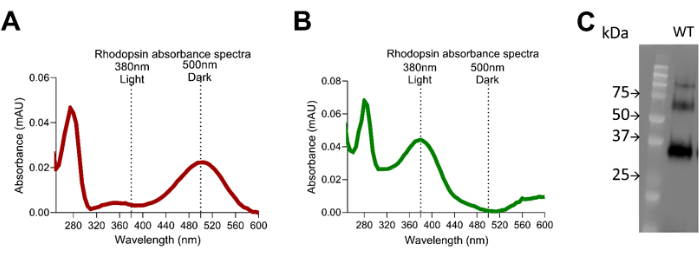
Figure 6: Rhodopsin absorbance spectra. (A) Dark adapted mice retina extract using 1D4 purified rhodopsin showing the 500 nm peak absorbance, and (B) light exposed retina extract using 1D4 purified rhodopsin showing the 380 nm peak absorbance. (C) Western blot using 1D4 rhodopsin antibody showing the quality of the rhodopsin purification and size distribution. Please click here to view a larger version of this figure.
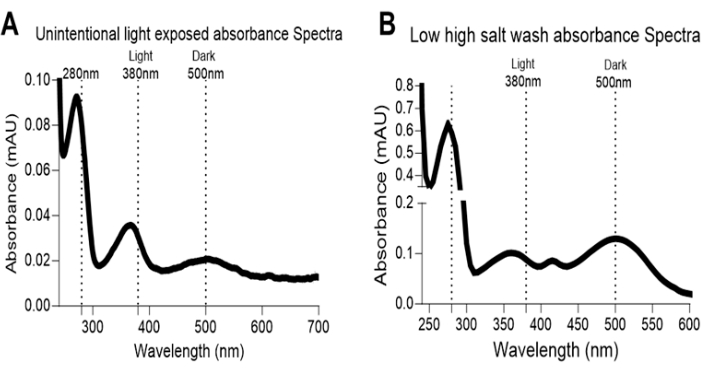
Figure 7: Rhodopsin absorbance spectra artifact. (A) Artifact in absorbance spectra of dark-adapted eye extract is shown with the absorbance peak at 380 nm; the unintentional light exposures from the instruments led indicators or failure of dark room door light blocker causes bleaching of rhodopsin protein. (B) Insufficient washes with high salt buffer cause accumulation of contaminates from RPE, lens, or other sources from eye extract, leading to high absorbance spectrum spread and artifacts. Please click here to view a larger version of this figure.
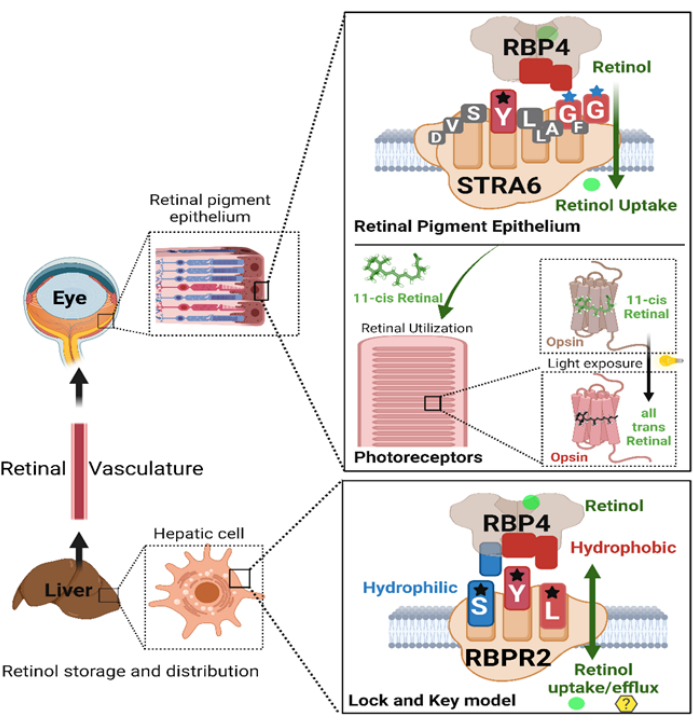
Figure 8: Schematic representation of the interactions between vitamin A membrane receptors (RBPR2 and STRA6) and Opsin protein with their respective ligands (RBP4 and 11-cis retinal) critical for generating the retinylidene protein in photoreceptors for visual function. Please click here to view a larger version of this figure.
Supplemental Figure S1: Intrinsic tryptophan fluorescence intensities. Preliminary evidence of interaction and change in the orientations of intrinsic tryptophan residues of RBP4 upon interacting with various concentrations of RBPR2 and STRA6 peptides. Please click here to download this File.
Supplemental Figure S2: RBPR2 and RBP4 protein-protein docking and interaction. Interface residues are visualized in PyMOL and the orientation of RBPR2 in the membrane is visualized by using the server tool https://opm.phar.umich.edu/. Please click here to download this File.
Supplemental Figure S3: Pipeline for in-silico analysis of the protein-protein docking interaction and possible contact sites. The analysis pipeline demonstrates the sequence of Python script run and PyMOL command line usage from steps 1 to 6, ensuring the reliability of acquiring the sites of interactions. Please click here to download this File.
Supplemental Figure S4: CD spectroscopy secondary structure analysis showing the composition of the RBP4, RBPR2, and STRA6 peptides. The pie graph shows the shares of secondary structures present in the molecule. Using the BeStSel Secondary Structure tool, Protein Fold Prediction by Circular Dichroism Spectroscopy (https://bestsel.elte.hu) was analyzed. Please click here to download this File.
