Acclimation to the behavior chamber involves three days of training: 2 days of group acclimation followed by 1 day of individual acclimation. However, because we could not distinguish individual zebrafish from one another, we were only able to collect data during individual acclimation. At this time, experimental animals (n = 30), conditioned using a shoal-based reward, took an average of 125.11 s to reach their first decision (Figure 2A) and an average of 725.34 s (12 min) to complete the entire individual acclimation task (Figure 2B). There was no significant side preference during acclimation (Figure 2C). The number of excluded fish was minimal as compared to other reward types (food) we had previously assessed in our laboratory (Figure 2C).
After acclimation, zebrafish began the acquisition phase. As fish were tested individually, we collected data from each fish on each of the three acquisition days. The fish were classified as either ‘high or low performers’, with ‘high performers’ responding faster and more accurately, despite all fish having the same previous exposure to the testing chamber. Only fish that selected the rewarded choice compartment in at least 6 of the 8 trials were classified as ‘high performers.’ Fish that did not meet this criterion were ‘low performers.’ High and low performing fish were housed separately to distinguish their performance in all subsequent trials. Interestingly, we observed that some fish changed category (i.e., were initially ‘low performers’, but became ‘high performers’) during the course of acquisition. In fact, the number of high performing animals increased each day, with more high performing fish on acquisition day 3 compared to day 1 (Figure 3A). By day 3, >50% of the fish had become ‘high performers.’ Further, initial choice latency for all fish across the three acquisition days (A1–A3) decreased, indicating an improved performance with each day of acquisition (Figure 3B). The same trend was also seen when only the high performing fish group was considered: by day 3, the time to first decision improved (became faster) (Figure 3C).
A discrimination ratio (rewarded trials/(reward + nonrewarded trials) was calculated for each acquisition trial block (average of two trials/fish) across the three acquisition days (A1–A3) for all experimental animals (n = 30) to determine how accurately the fish were solving (acquiring) the discrimination task (i.e., going to the rewarded side of the tank). This ratio revealed that the percentage of fish moving to the rewarded side during each trial increased daily (i.e., across trial blocks on each individual day) and overall (i.e., across the three acquisition days) resulting in all fish performing above chance by the end of acquisition (dotted line represented on the graph; (Figure 4A) and indicating that the fish had learned the discrimination task.
Following the acquisition of discrimination learning, we tested how long zebrafish would remember the task. To do that, tested zebrafish remained in holding tanks for 8 weeks. After this time, fish were tested on a reversal task that lasted 3 days (R1–R3). We found that the fish demonstrated strong reversal behavior and increased discrimination over the three days of reversal (Figure 4B), indicating they were able to (1) remember the relationship between the color of the tank and the reward and (2) inhibit what they had previously learned during acquisition and learn the reversal/opposite paradigm. As shown in Figure 4B, zebrafish initially went to the non-rewarded side of the tank, as indicated by the discrimination ratio being below chance during initial trails on reversal day 1. However, by the end of R1, performance increased to greater than chance, a result that was maintained on R2 and R3, with the highest discrimination ratio scores observed on R3. Taken together, these data show that naïve experimental animals are capable of solving the discrimination task, even though the initial behavior was acquired 8 weeks earlier, without any additional training in between behavioral sessions.
The 3-chamber choice paradigm can also be applied to examination of disease complications. In our study with hyperglycemic zebrafish, acclimation and acquisition were as described, and reversal was tested following either 4 or 8 weeks of hyperglycemia. Hyperglycemia was induced with an alternate immersion protocol (McCarthy et al., 2020 – this issue), so that training occurred every other day, on days after zebrafish had been in test solutions for 24 h. During acquisition, there was a main effect of training day on discrimination ratio (F (2, 239) = 4.457, p = 0.012; Figure 5A), with the ratio on A1 being significantly lower than on A3 (p = 0.010), indicating that the fish improved their choice accuracy over time. During reversal, there was a significant main effect of treatment (F (2, 326) = 3.057, p = 0.048), but no other significant main effects or interactions (training day: (F (2, 326) = 1.602, p = 0.203); training day x treatment: (F (4, 326) = 0.661, p = 0.620); Figure 5A). The response of glucose-treated animals was significantly reduced compared to the water-treated animals (p = 0.037), but there were no other significant differences (control v. mannitol: p = 0.387; mannitol v. glucose: p = 0.524), suggesting a glucose-specific effect. After 8 weeks of hyperglycemia, no statistical differences were noted in discrimination ratios across acquisition training (F (2,263) = 2.909, p = 0.056; Figure 5B). However, there were significant main effects of both training day (F (2, 189) = 4.721, p = 0.010) and treatment (F (2, 189) = 7.940, p = 0.000) on reversal, but no significant interaction (training day * treatment = F (4, 189) = 0.869, p = 0.484). Subsequent least significant difference (LSD) pairwise comparisons identified significant differences between R1 and R3 (p = 0.022) and between R2 and R3 (p = 0.003). LSD pairwise comparisons also revealed significant differences between the water treatment group and both glucose and mannitol treatment groups (water v. mannitol: p = 0.008; water v. glucose: 0.000); however, the glucose and mannitol groups were not significantly different from one another (p = 0.265), suggesting that these differences in discrimination ratio may be due to osmotic effects.

Figure 1: Three-chamber choice testing chamber and behavior setup. (A) 3-chamber schematic. Experimental animals were restricted to the central starting chamber for 2 min and then allowed access to either side of the tank at the start of a trial. To do so, the top half of each of the two partitions were raised to create a 10 cm space for fish to cross into either choice compartment. (B,E) Acclimation was performed using a beige background and shoal of conspecifics as the reward. (C,F) Acquisition was performed using black and white backgrounds on the choice chambers; reward was only located on one side of the chamber. (G) Reversal was performed using black and white backgrounds on the choice compartments; reward was only available on the opposite side of the chamber (vs. acquisition). (H) Up-close image of shoal tank submerged in one of the choice compartments. Please click here to view a larger version of this figure.
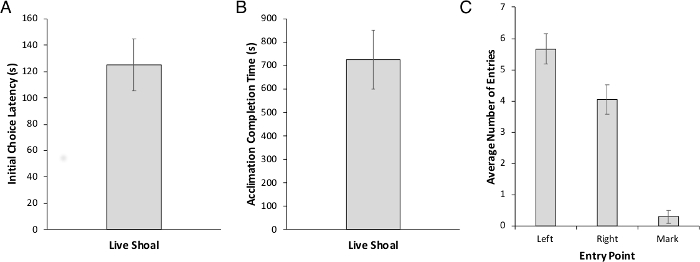
Figure 2: Latency and number of marked trials during individual acclimation. (A) Choice latency of first decision. (B) The total amount of time to complete individual acclimation. (C) The number of entries to the left and right side are not different, indicating no inherent side preference prior to the start of acquisition. We also report the total number of mark trials during individual acclimation. Values are reported as Mean ± SEM. Please click here to view a larger version of this figure.
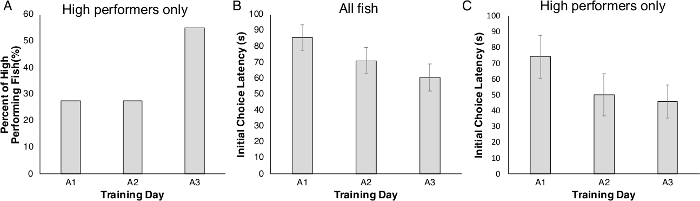
Figure 3: The percentage of high performing fish and the initial choice latency among all fish and high performing fish across the three acquisition days. (A) High performing fish moved from the central chamber to the rewarded side of the chamber in at least 6 out of 8 trials each acquisition day (A1–A3). (B) Across three days of acquisition training (A1–A3), overall initial choice latency decreased; a trend also evident in high performing fish (C). Values are reported as Mean ± SEM. Please click here to view a larger version of this figure.
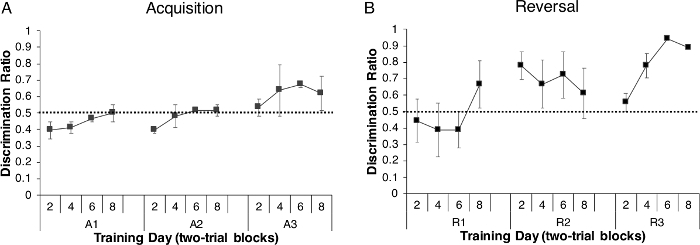
Figure 4: Discrimination performance during acquisition and reversal trials. (A) Discrimination ratio (rewarded trials/(reward + nonrewarded trials) of fish across acquisition days (A1–A3) and (B) during reversal learning 8 weeks later. Reversal was also assessed for 3 days (R1–R3). For both tasks, each fish had to complete 8 trials, and results are presented in two-trial blocks (2, 4, 6, 8). Correct responses during both acquisition and reversal increased with time, with a faster response observed during reversal, indicating the fish learned and remembered the task. Values are reported as Mean ± SEM. The dotted line represents chance. Please click here to view a larger version of this figure.
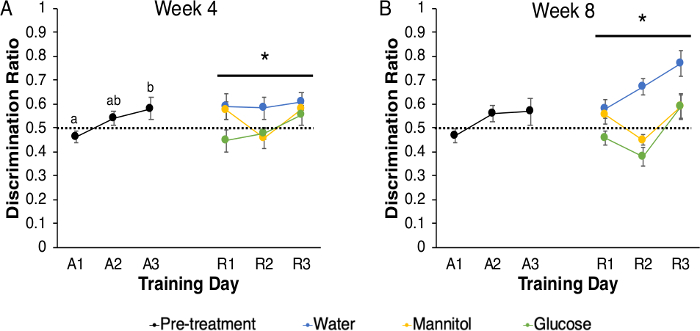
Figure 5: Acquisition and Reversal of hyperglycemic zebrafish using the 3-chamber choice behavioral task. (A) Prior to treatment, naïve zebrafish acquired the 3-chamber choice behavioral task across three days of behavioral training (acquisition, A1–A3). There was a significant difference between discrimination ratios on A1 and A3 indicating learning took place (p = 0.012). Following 4 weeks of treatment (colored symbols) there was a significant effect of treatment (p = 0.048), with glucose-treated animals displaying significantly reduced discrimination ratios compared to water-treated animals (p = 0.037). (B) In a separate experiment, behavior before and after 8 weeks of hyperglycemia was assessed. Despite the steady increase in performance across each acquisition day, there were no significant differences discrimination ratio across A1–A3. However, after 8 weeks of treatment (colored symbols), there was a main effect of treatment (p < 0.001) and an individual main effect of training day (p = 0.010). Post-hoc analyses revealed a significant difference between the water-treated group and both mannitol- and glucose-treated groups, suggesting an osmotic effect (water v. mannitol: p = 0.008; water v. glucose: p < 0.001). * denotes a significant main effect. Data points represent group mean ± SEM, and data points with different letters are significantly different from one another. The dotted line represents chance. Please click here to view a larger version of this figure.
