本研究では、5つのアポCIII KOウサギをZFN技術を用いて生成した。以前のGTTウサギの生産の唯一の報告書はまた、ZFN技術18を使用していました。本研究では、ZFNを効果的にウサギの遺伝子を標的にすることが有用であることを確認した。
本研究では、ウサギの前のZFNレポート(30.8%、52分の16)18に匹敵する、それらが報告された遺伝子組換え率(正キット/合計生まれ)is23.8%(5月21日)、ゼブラフィッシュ19、マウス20、21匹のラットおよびブタ17を含む他の動物種において、ZFNを用いた。この速度は、実際には、通常、5〜20%の範囲に多くの従来のトランスジェニックウサギの生産試験のレートよりも高い。例えば、従来のDNAのマイクロインジェクションを介した製品アポCIIIトランスジェニックウサギに努め、鼎ら (5.6%)22生まれの54匹のうち3つの肯定的な創業者を得た。
これまでの知見と一致して、ターゲットのサイトでの変異は、(5 KOのウサギで1から21までの範囲)の塩基数が異なるから欠失または挿入を含む、可変である。創始者動物の特定の配列情報に基づいて、少なくとも4つのことが予想される( すなわち 。Δ1、+1、+1、またはΔ20変異を含有するもの)はアポCIIIの機能喪失を有するであろう。この変異が唯一の原因の7アミノ酸の喪失ではなく、読み枠シフトすることが予測されるなどの動物、R-16Δ21では、機能の損失が表示されない場合があります。最終的に、表現型アッセイは、これらの創始動物が本当にアポCIII機能の喪失を表示するかどうかを決定するために必要である。
本研究で生成された5 KOウサギはいずれも二対立遺伝子の修正が含まれていない。興味深いことに、これはまた、以前のZFNウサギのレポートの場合である。これとは対照的に、ZFNはα1、3 – ガラクトシルをターゲットとした場合は、豚に適用された細胞は、単一の対立遺伝子を標的化頻度は、突然変異の約3分の1は23ノックアウト対立遺伝子、シングルステップの作成 を、7から46までパーセントの範囲であった。 TALENはブタ線維芽細胞に適用したときにこれと一致して、対立遺伝子改変は、合計14個の陽性クローンの約15〜40%に認められた。なお、本研究では二対立遺伝子のKOウサギを生成する失敗がターゲットとヌクレアーゼベースの遺伝子のような能力のための種差( すなわちウサギ対豚)を指示する可能性がある。なお、このプロジェクトで生成KOウサギの数が比較的少ないため、これは単にであることがより可能性が高い。我々は、ZFNは、ZFNベースのGTTウサギ生産の別の潜在的な利点として考慮されるべきであるウサギにbialleic突然変異を発生させることができると信じています。さらなる実験は、ウサギにおけるZFNのような能力を確認するために必要とされる。
結論として、本研究では実証するZアプローチを対象とした、FNベースの遺伝子がノックアウトウサギを生産するのに効果的である。特に、我々は、23.8%の満足ジェニック率は5アポCIIIのKOウサギを生成しました。これらの動物は、対応するマウスモデルよりも、ヒトにおける脂質代謝に対するこのタンパク質の役割についてのより意味のある情報を提供すると考えられている。我々は、ZFNベースの遺伝子ターゲティング、ならびにTALENやRGENなど他のヌクレアーゼベースの技術を予測、nonmurine動物で有意に様々なヒトの病気を研究するための新しい動物モデルの開発を促進します。
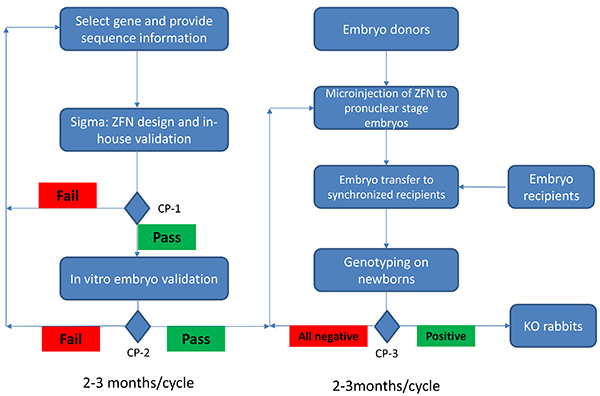
図1。 ZFN技術を用いてノックアウトウサギの産生のチャートを流す 。 ZFN技術を使用してのKOウサギの生産は、目的の遺伝子の選択から始まります。配列情報が提供され、ZFNセットが設計されている首題社内での検証に基づく酵母ected。チェックポイント(CP)1(CP-1)では、社内検証(内部陽性対照の50%以上)を通過したもののみセットが選択のためにユーザに提供される。まだセットはCP-1に合格しない場合は、追加の配列は、効果的なZFNセットを設計するために必要とされ得る。 in vitroでの胚の検証は、選択したZFNセットが選択したサイト(CP-2)に突然変異を誘発することができることを確認するために続いている。それは( インビトロ胚で> = 10%の突然変異率)CP-2に合格した場合、ZFNセット(S)がのみ使用されます。 CP-2での失敗は、ZFNセットの再設計が必要になります。胚ドナーが準備され、前核期胚を検証ZFNセットをマイクロインジェクションされます。これらのマイクロインジェクションした胚は、同期化された胚の受信者に転送されます。 1ヶ月の妊娠期間の後、新生児は(CP-3)遺伝子型を決定します。新生児のいずれも陽性でなかった場合、追加のマイクロインジェクションが行われる。それは、nを仮定し、CP-2に、プロジェクトの開始から2〜3ヶ月かかりますプロセス中のO障害。それは、CP-3へのCP-2からさらに2〜3ヶ月かかります。従って、ZFN技術を使用して4-6ヶ月の時間枠内でノックアウトウサギを生成することができる。 拡大画像を表示するには、ここをクリックしてください 。
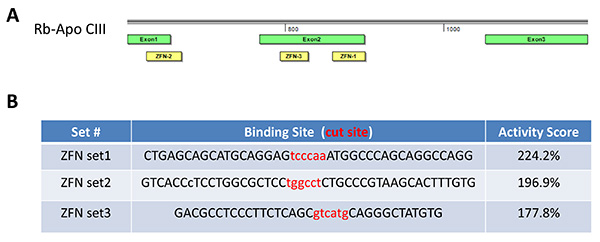
図2。 ZFNデザイン 。三ZFNは(セット1、2、および3)を設定するエキソン1またはウサギアポCIIIのエクソン2(A)上の異なる配列を標的とするように設計した。すべての3つのセットは、ZFN活性を決定するために、酵母MEL-1レポーターアッセイに供した。製造業者のプロトコールに従って、製造業者の内部陽性対照のそれの> 50%の活性を示すのZFNは、 インビトロおよびインビボのゲノム編集のための実験において有用であるとみなされる。ZFN活動は、したがって、1本研究では、選択した設定、セット1のための224.2パーセント、セット2のための196.9パーセントとセット3(B)のための177.8パーセントだった。 拡大画像を表示するには、ここをクリックしてください 。
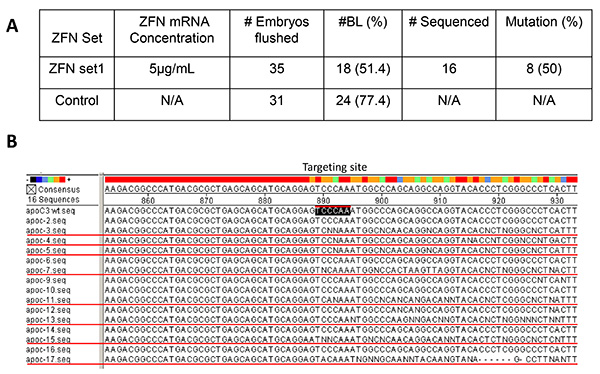
ZFNの図3。胚の体外検証 。セット1のmRNA(5μgの個/ ml)を、35前核期ウサギ胚(A)の細胞質にマイクロインジェクションした。マイクロインジェクション治療た(n = 31)なしでフラッシュした胚を対照群として使用した。 BLの展開率は、対照群で77.4%であった。顕微注入群では、BL率は51.4%だった。 PCR配列を受け16 BLのうち、8(50%)が陽性と確認された、セット1を示すことはタージェで突然変異を誘発するための効果的なZFNセットですティンサイト(B、apoc3wt.seq、ブラックボックス化)ウサギにおけるアポCIIIの。突然変異を含有するが、BLとBL#のAPOC-3、4、5、7、11、13、15、および17である(B、赤下線)。残りのBLは(#のAPOC-2、6、9、10、12、14および16)は、標的部位での変異を持っていません。 拡大画像を表示するには、ここをクリックしてください 。

図4。アポCIIIのKOウサギの生産 。 ZFNセット1のmRNAをマイクロインジェクションは百四〇から五胚は7偽妊娠レシピエントウサギ(A)に移した。 ZFNマイクロインジェクションすることなく新たにフラッシュした胚た(n = 75)(n = 6)は、対照群としてレシピエントに移した。実験群では、全期間キット/総胚として計算期間率は14である.5%(21/145)、速度は対照群では(22/75)29.3%であるのに対し、(A)。顕微注入群で生まれた21のキットのうち、5個(R1、R9、R11、R12およびR16)を、PCR配列決定(B)後に陽性KOキットとして同定された。標的サイトのインデルは2つの挿入(R9とR11の両方のための1)と(それぞれΔ1、Δ20、R1、R12とR16のためのΔ21)3欠失を含む。 拡大画像を表示するには、ここをクリックしてください 。
