In the present work, five ApoCIII KO rabbits were generated using the ZFN technology. Previously the only report of the production of GTT rabbits also used the ZFN technology18. The present work confirmed that ZFN is useful to effectively target genes in rabbits.
In the present study, the transgenic rate (positive kits/total born) is23.8% (5/21), which is comparable to that of the previous ZFN report in rabbits (30.8%, 16/52)18, and those reported of using ZFN in other animal species, including zebra fish19, mice20, rats21 and pigs17. This rate is in fact higher than the rates of many conventional transgenic rabbit production studies, which normally fall in the range of 5-20%. For example, in an effort to product ApoCIII transgenic rabbits via conventional DNA microinjection, Ding et al. obtained 3 positive founders out of 54 pups born (5.6%)22.
Consistent with previous findings, the mutations at the targeting sites are variable, including deletions or insertions consisting different number of bases (ranging from 1-21 in the five KO rabbits). Based on the specific sequence information of the founder animals, it is predicted that at least four (i.e. those containing Δ1, +1, +1, or Δ20 mutations) would have function loss of ApoCIII. Animal R-16 with Δ21 may not display functional loss, as this mutation is predicted to only cause loss of seven amino acids, but not a reading frame shift. Ultimately, phenotype assays are necessary to determine if these founder animals truly display the loss of ApoCIII functions.
None of the five KO rabbits generated in the present study contain biallelic modifications. Interestingly, this is also the case in the previous ZFN rabbit report. In contrast, when ZFNs targeting α1,3-galactosyltransferase were applied to pig cells, the frequency of targeting a single allele ranged from 7-46%, with approximately one-third of mutations creating single-step biallelic knockouts 23. Consistent with this, when TALEN was applied to pig fibroblast cells, the biallelic modifications were found in approximately 15-40% of the total positive clones14. It is possible that the failure to generate biallelic KO rabbits in the present study may indicate a species difference (i.e. rabbits vs. pigs) for such capacity of nuclease based gene targeting. It is, however, more likely that this is simply because the number of KO rabbits generated in this project is relatively small. We believe that ZFN is capable of generating bialleic mutations in rabbits, which should be considered as another potential advantage of ZFN based GTT rabbit production. Further experiments are needed to confirm such capacity of ZFN in rabbits.
In conclusion, the present work demonstrates that ZFN based gene targeting approach is effective in producing KO rabbits. In particular, we generated five ApoCIII KO rabbits with a satisfactory transgenic rate of 23.8%. These animals are believed to provide more meaningful information about this protein’s role on lipid metabolisms in humans than the corresponding mouse models. We predict ZFN based gene targeting, as well as other nuclease based technologies such as TALEN and RGEN, in nonmurine animals will significantly facilitate the development of novel animal models to study various human diseases.
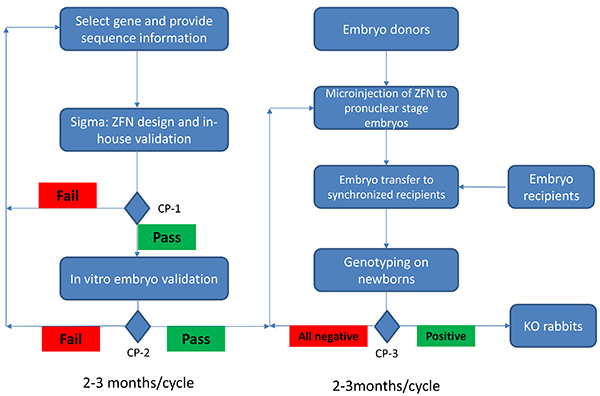
Figure 1. Flow chart of production of knockout rabbits using ZFN technology. Production of KO rabbits using ZFN technology starts with the selection of the gene of interest. The sequence information is provided, ZFN sets are designed, and subjected to yeast based in-house validation. At check point (CP) 1 (CP-1), only those sets passed in-house validation (50% or higher of the internal positive control) will be provided to the users for selection. If no sets pass CP-1, additional sequence may be needed for designing effective ZFN sets. An in vitro embryo validation is followed to ensure the selected ZFN set can induce mutations at selected sites (CP-2). ZFN set(s) will only be used if it passes CP-2 (>=10% mutation rates in in vitro embryos). Failure at CP-2 will require a redesign of the ZFN sets. Embryo donors will be prepared and pronuclear stage embryos will be microinjected with the validated ZFN sets. These microinjected embryos will be transferred to synchronized embryo recipients. After one month gestation period, newborns will be genotyped (CP-3). If none of the newborns are positive, additional microinjection will be performed. It takes 2-3 months from start of the project to CP-2, assuming no failure during the process. It takes additional 2-3 months from CP-2 to CP-3. Therefore it is possible to generate a knockout rabbit in a 4-6 month time frame using the ZFN technology. Click here to view larger image.
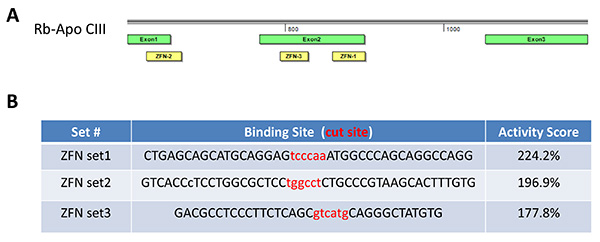
Figure 2. ZFN design. Three ZFN sets (Set 1, 2, and 3) were designed targeting different sequences on Exon 1 or Exon 2 of rabbit ApoCIII (A). All three sets were subjected to the yeast MEL-1 reporter assay to determine the ZFN activities. According to the manufacture’s protocols, ZFNs that show >50% activities of that of the manufacture’s internal positive control are regarded as useful for in vitro and in vivo genome editing experiments. The ZFN activities were 224.2% for Set 1, 196.9% for Set 2 and 177.8% for Set 3 (B), therefore Set 1 was selected in the present study. Click here to view larger image.
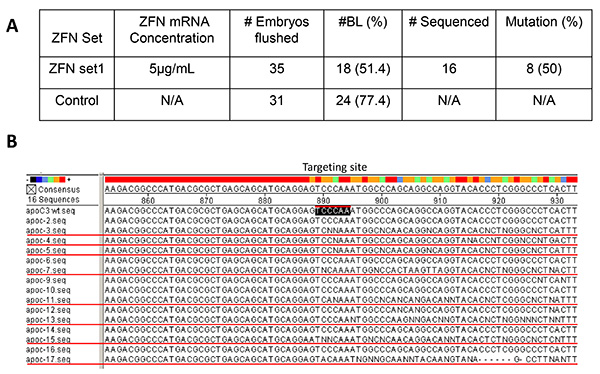
Figure 3. In vitro embryo validation of ZFN. The mRNA (5 μg/ml) of Set 1 was microinjected to the cytoplasm of 35 pronuclear stage rabbit embryos (A). Flushed embryos without microinjection treatment (n=31) were used as the control group. The BL development rate was 77.4% in the control group. In the microinjected group, the BL rate was 51.4%. Of 16 BLs subjected to PCR sequencing, 8 (50%) were identified as positive, indicating Set 1 is an effective ZFN set to induce mutations at the targeting site (B, apoc3wt.seq, black-boxed) of ApoCIII in rabbits. The BLs containing mutations are BL#apoc-3, 4, 5, 7, 11, 13, 15, and 17 (B, red underlined). The remaining BLs (#apoc-2, 6, 9, 10, 12, 14 and 16) do not have mutations at the targeting site. Click here to view larger image.

Figure 4. Production of ApoCIII KO rabbits. One hundred and forty-five embryos microinjected with ZFN Set 1 mRNAs were transferred to 7 pseudo-pregnant recipient rabbits (A). Freshly flushed embryos (n=75) without ZFN microinjection were transferred to recipients (n=6) as the control group. The term rate calculated as total term kits/total embryos in experiment group is 14.5% (21/145), whereas the rate is 29.3% (22/75) in the control group (A). Out of 21 kits born in the microinjected group, five (R1, R9, R11, R12 and R16) were identified as positive KO kits after PCR sequencing (B). The indels at the targeting site include two insertions (+1 for both R9 and R11) and three deletions (Δ1, Δ 20, Δ 21 for R1, R12 and R16, respectively). Click here to view larger image.
