The images obtained with this methodology have an intrinsic low background level due to the very limited number of harmonophores present in biological tissues, which is one of the significant advantages of the method.
When the fibers of the corpus callosum are imaged, fiber-like short structures and rounded elements can be consistently found in the taiep brain (Figure 3B), while the corpus callosum of the control brain shows a much more heterogeneous and isotropic signal throughout the brain region (Figure 3A). The origin of the differential signal lies specifically in the second harmonic generation phenomenon, since adding the narrow bandpass filter only decreases the non-specific signal intensity from the control images (Figure 3C–D) while selectively removing this low, diffuse signal from around the soma-like and the short, elongated structures in the taiep images, which always generate intense SH light (Figure 3E–F).
The other structure analyzed, the cerebellar white matter, gives comparable results. Specifically, while in control tissue there is an almost complete absence of SH signal (Figure 4B), and the Purkinje cells are barely visible when using the short-pass filter, the elongated and rounded structures persist in the SH image from the taiep tissue (Figure 4D).
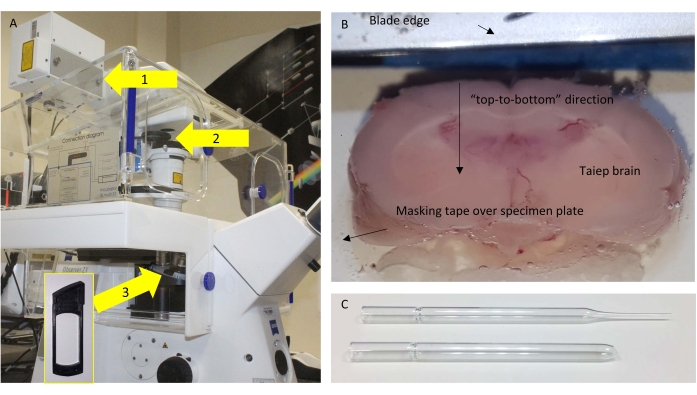
Figure 1: Microscope and sectioning. (A) Schematic of the microscope used, with relevant components highlighted. Arrows: 1 = NDD port where the SP485 is located close to the detector; 2 = removable frames where the BP405 is placed; 3 = position under the objective where the half-wave plate (HWP) is placed for control experiments. The inset shows the frame where the HWP could be inserted. (B) Top view of the vibratome buffer tray with the glued brain ready to be fine-sectioned. (C) The original (top) and modified (bottom) Pasteur pipette used to transfer the sections. Please click here to view a larger version of this figure.
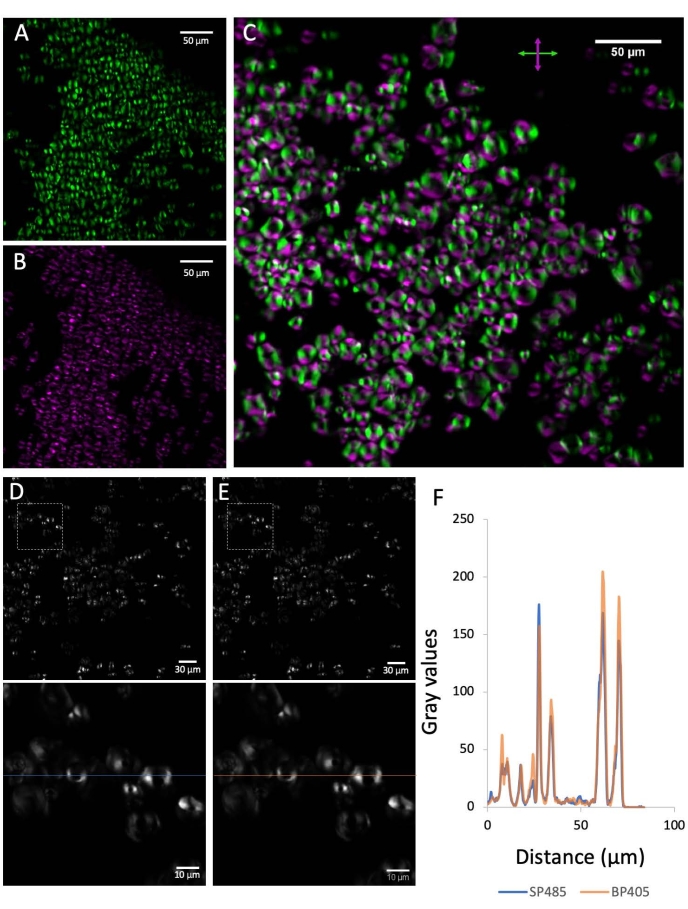
Figure 2: Preliminary controls. (A) The SHG signal emitted by corn starch grains shows a predominant orientation that coincides with the oscillation direction of the laser (horizontal in this case). (B) After inserting a half-wave plate with its fast axis oriented at 45° with respect to the horizontal, the signal is rotated by 90°. (C) Merging of the signal before and after the half-wave plate insertion. (D) The signal below 485 nm emitted from the corn starch grains. (E) The SHG signal from the starch grains detected at 405/10 nm. (F) Graph showing a comparison of the two signals corresponding to the line-scans shown in the zoomed inserts of D and E. The SHG filter used causes negligible signal loss. Please click here to view a larger version of this figure.
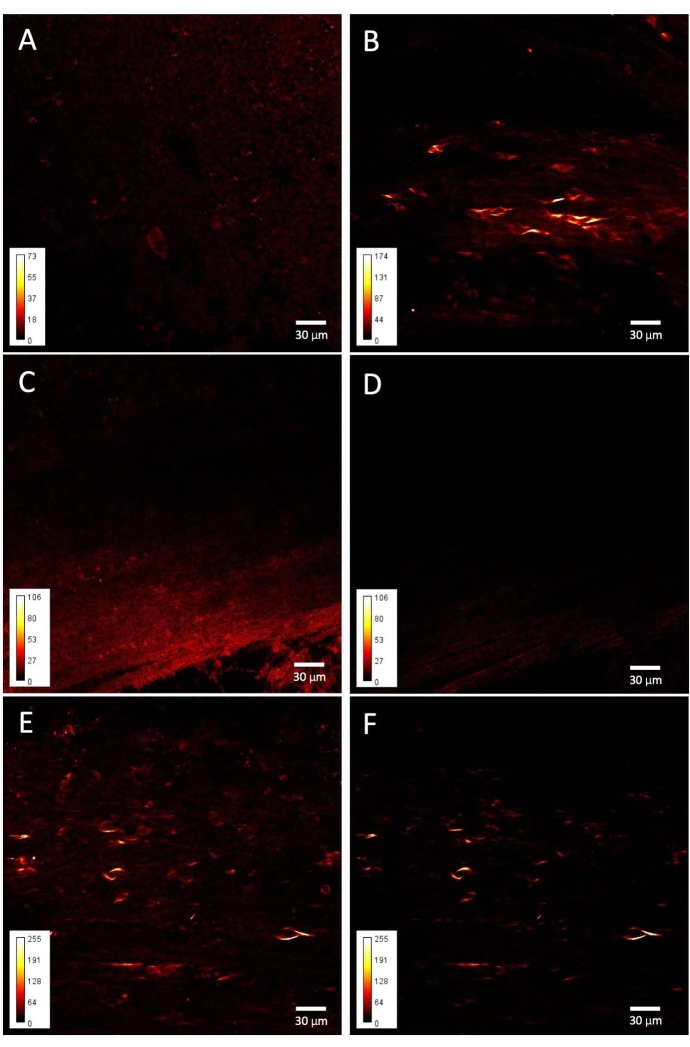
Figure 3: SHG images from taiep and WT corpus callosum. Representative examples of (A) WT and (B) taiep signals obtained from the corpus callosum. (C) SP485 filtered image from an WT corpus callosum. (D) BP405 filtered image from the same sample as in C. (E) SP485 filtered image from a taiep corpus callosum. (F) BP405 filtered image of the same sample as in E. Please click here to view a larger version of this figure.
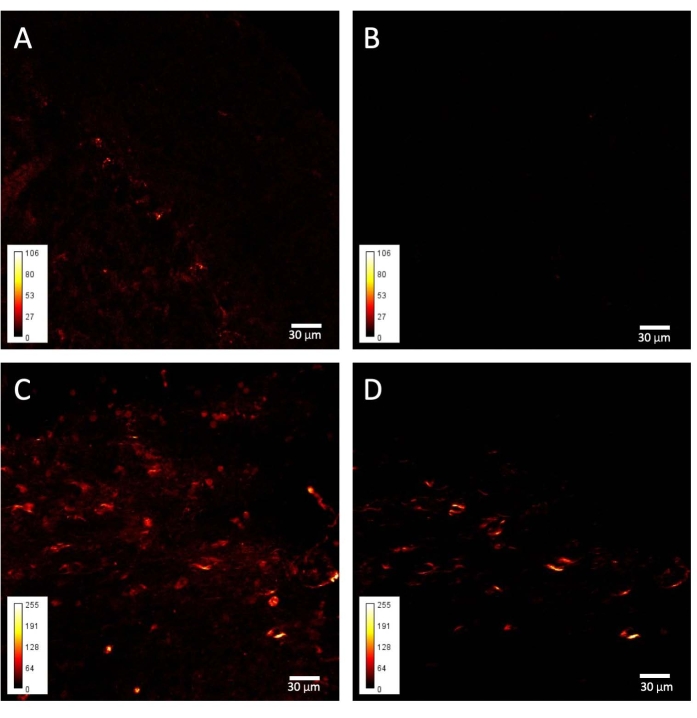
Figure 4: SHG images from taiep and WT cerebellums. Representative examples of (A–B) WT and (C–D) taiep signals obtained from cerebellar folia. (A) An SP485 filtered image from a WT folium; only some Purkinje cells are visible. (B) A BP405 filtered image from the same sample as in A. (C) An SP485 filtered image from a taiep folium. (D) A BP405 filtered image of the same sample as in C. Please click here to view a larger version of this figure.
Supplementary Figure 1: Examples of artifacts. (A) Artifact due to drying of the sample. (B) Artifact due to excessive exposure. Scale bars: 30 µm. Please click here to download this File.
Supplementary File 1: Köhler alignment.The file presents the steps for performing the Köhler alignment. Please click here to download this File.
Supplementary File 2: ZEN software steps for SHG image acquisition. Please click here to download this File.
