생체 분자의 구조를 결정하는 것은 그 기능을 이해하기위한 중요한 전제 조건이다. 구조 결정을위한 두 개의 잘 확립 된 방법은 극저온 전자 현미경 및 X 선 결정학 (1, 2)이다. 오늘, 두 가지 방법은 옹스트롬 수준 아래로 해상도를 가진 고해상도 구조 정보를 제공합니다. 이러한 두 가지 방법은 단백질 복합체 같은 큰 생 분자의 구조를 규명하기 위해 광범위하게 사용되고있다. 기존의 방법은 지속적 지난 수십 년에 걸쳐 개선되어 왔지만, 큰 동적 과도 복합체 3을 조사하는 경우, 생물학적 구조의 복잡성은 여전히, 특히 구조 생물학에 중요한 도전을 제기.
고분자 복합체의 역학, 특히 구조 – 기능 관계를 연구하기 위해서, 단일 분자 방법론 잠을ided 유용한 정보 4. 여러 새로운 전략 구조 및 동적 정보를 획득하는 방법에 직교 제공 개발되었다. 예 고속 AFM (5) 기계적 조작 6 지역화 형광 현미경 (7)뿐만 아니라, 단일 분자 포스터 공명 에너지 전이 (smFRET) (8, 9)이다. FRET 매우 이른 이후에 의한 생체 거대 분자 (10)의 길이 규모의 거리에 의존하는 분자자를 지칭되었다.
smFRET의 한 특히 흥미로운 애플리케이션은 추론 smFRET 측정으로부터 얻어진 거리 정보를 사용하는 정보는 11, 12, 13, 14, 15의 구조 </sup> 16, 17, 18, 19, 20, 21, 22, 23. smFRET 인해 높은 시간 분해능으로, 단백질 구조의 이동 부분의 위치는 국소 수있다. 그러나, 염료 분자에 대한 smFRET 데이터 중요한 보정 파라미터로부터 정량적 정보를 추출하기 위해 측정을 24 동안 결정되어야한다. 이들 보정 계수로, FRET 효율 E FRET은 공식을 사용하여 계산 될 수있다
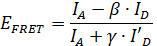 ,
,
어디 A와 I D </sUB는> 각각 공여체 및 수용체 분자의 형광 강도 (도 2 참조)이다. β-인자 수용체 채널에 크로스 – 토크에 대한 도너 발광 누설 계정에 의해 계산된다

나는 'A와 나'D 경우 기증자의 형광 강도와 수용체 분자의 사진 표백 후 수용체 분자이다.
γ 인자는 두 채널의 검출 상대 효율의 차이뿐만 아니라 도너의 형광 양자 수율 셉터 염료의 차이를 보정한다. 그것은으로 모든 개인 시간 추적 계산한다
<p class="jove_content" fo:keep-together.within-page="1" fo:text-align="center" style = "텍스트 정렬 : 센터;">
이 설명은 종종 중요뿐만 아니라 보정 될 필요 셉터 분자의 직접적인 자극을 무시한다는 참고. 이러한 보정 인자를 결정하는 것은 광 신체적 변화 및 구조 역학 구별하기 위해 교류 방식 (25)에 도너뿐만 아니라 수용체 모두를 여기 시키는데 유용하다.
정량적 smFRET 효율뿐만 아니라 양적 구조 정보를 얻을뿐만 아니라 위해, 나노 포지셔닝 시스템 (NPS)는 2008 (26)에 도입 하였다. 이름은 위성 기반의 위성 위치 확인 시스템 (GPS)과의 유사성을 기준으로 선택되었다. 국민 연금은 biomacromolecular 단지에서 알 수없는 염료 위치의 현지화를 위해 smFRET 및 X 선 결정학 데이터를 결합하는 하이브리드 기술이다. 는 Crystal 구조는 기준 프레임으로서 작용하고 smFRET 결과를 알 수없는 위치 형광 (안테나)과 결정 구조 (위성)로부터 공지 된 위치 사이의 거리 정보를 획득하기 위해 사용된다. 연속적인 실험에서 안테나와 여러 개의 위성들 사이의 거리가 측정되고, 상기 안테나의 위치는 베이지안 파라미터 추정에 기초하여 통계적으로 엄격한 분석 기법에 의해 결정된다. 결과적으로, 상기 안테나의 위치를 가장 가능성뿐만 계산되지만 완전한 3D 불확실성 분포 소위 후방은 신뢰할 볼륨 시각화. 더욱이, NPS smFRET 완전한 네트워크 (27)의 분석을 허용하도록 확장되었다.
NPS는 진핵 전사 중요한 질문 상류 DNA, 비 – 주형 DNA 및 RNA 중합 효소 II 신장 공동 내의 초기의 mRNA 즉 과정들을 해결하기 위하여 사용되어왔다또한 전사 개시의 효과를 보여주는 mplex 12, 28, 26 요인 29 오픈 프로모터의 동적 구조는 복잡. 더욱이, NPS는 전사 연장 인자 Spt4 / 5 (31)와 동일한 위치에 경쟁적으로 결합하는 전사 개시 인자 TFE의 위치는 고세균 RNA 폴리머 라제 오픈 착체 (30)의 특히 구조를 규명 하였다.
이후 smFRET 기반 구조 방식의 수는 15, 18, 21, 23을 발표 하였다. 다른 smFRET 기반 구조 방법을 비교하면,이 방법의 정밀도 명백한 색소 모델의 특정 선택에 크게 의존하는 것이 분명해진다. 하나는주의해야염료 분자는 지역 환경에 따라 서로 다른 공간과 배 양성 문제가 발생할 수 있습니다.
이를 위해, 빠른 NPS는 32 도입되었다. 고속 NPS 크게 계산 시간을 감소시키는 향상된 샘플링 알고리즘을 사용한다. 또한, 고속 NPS는 하나의 구조 분석을 수행하고 각 염료 분자의 사용자는 다음에 설명한다 다섯 가지 염료 모델 세트를 선택할 수 있습니다. 고전 불리는 가장 보수적 모델 염료는 하나이지만 알려지지 않은 위치를 차지하는 것으로 가정한다. 이 위치에서, 형광 물질은 크기가 각각의 (시간에 따른) 형광 이방성 결정하는 콘에서 자유롭게 회전 할 수있다. 원뿔의 방향으로 측정 된 거리 smFRET 효율 변환시 큰 불확실성에 이르게하는 공지되지 않는다. 그것은 다른 염료 모드에 비해 작은 정밀도로 이끌 것이기 때문에 이러한 점에서, 모델은, 보수적LS. 매우 짧은 거리를해야 띄게 잘못된 위치 결정에 클래식 모델 리드에 의해 가정. 전형적인 smFRET 값의 경우, 정확한 위치는 항상 비교적 큰 신뢰할 수있는 볼륨에 묶여 있습니다.
보다 정밀 바람직하므로 그러나, 개발 및 정밀도를 향상시키기 위해 도움이 될 다른 염료 모델을 시험하는 것이 중요하다. 염료 고유의 형광 수명보다 빠르게 회전하면, 소위 ISO 모델을 적용 할 수있다. 여기서, 2 κ 배향 계수 (특성 등방성 포스터 반경을 계산하기 위해 필요한  ) 2/3로 설정됩니다. 전형적인 모델 (32)에서와 비교하여 그 결과 계산 신뢰할 볼륨은 작은 크기의 약 2 명령이다. 빠른 reori뿐만 있도록 형광이있는 환경에서 발견되는 경우에모든 액세스 볼륨 이상이 정해져있는,하지만 또한 빠른 동작은 meanpos 이소 모델이 사용되어야한다. 이 모델에서, 상기 염료는 효과적으로 공간적 평균화 다항식 거리 변환 (15)에 의해 차지되는 평균 하나의 위치를 차지한다. 예를 들어, (일반적으로 소수성) 염료는 친수성 영역, 예를 들면, DNA에 연결되어있는 경우,이 모델이 적용됩니다. meanpos 이소 모델의 적용은 약 2의 팩터에 의해 신뢰할 볼륨의 크기를 더욱 감소 리드. 그러나, 단백질에 결합 염료는 입체적 액세스 볼륨 (AV)의 몇몇 소수성 패치에 가역적으로 결합 할 수 있습니다. 순간적으로이 지역 사이에서 전환하지만, 한 지역이 자유 회전을 겪게하고 내 빠른 현지화 움직임이 가장 VAR-meanpos 이소 모델에 의해 설명되는 형광. 비슷한 상황에있는 염료는 모델이 적용되는 VAR-meanpos을 자유롭게 회전 할 수 없습니다. 더 라 이 모델에 대한 etails는 우리의 최근 출판물 (32)에서 찾을 수 있습니다.
) 2/3로 설정됩니다. 전형적인 모델 (32)에서와 비교하여 그 결과 계산 신뢰할 볼륨은 작은 크기의 약 2 명령이다. 빠른 reori뿐만 있도록 형광이있는 환경에서 발견되는 경우에모든 액세스 볼륨 이상이 정해져있는,하지만 또한 빠른 동작은 meanpos 이소 모델이 사용되어야한다. 이 모델에서, 상기 염료는 효과적으로 공간적 평균화 다항식 거리 변환 (15)에 의해 차지되는 평균 하나의 위치를 차지한다. 예를 들어, (일반적으로 소수성) 염료는 친수성 영역, 예를 들면, DNA에 연결되어있는 경우,이 모델이 적용됩니다. meanpos 이소 모델의 적용은 약 2의 팩터에 의해 신뢰할 볼륨의 크기를 더욱 감소 리드. 그러나, 단백질에 결합 염료는 입체적 액세스 볼륨 (AV)의 몇몇 소수성 패치에 가역적으로 결합 할 수 있습니다. 순간적으로이 지역 사이에서 전환하지만, 한 지역이 자유 회전을 겪게하고 내 빠른 현지화 움직임이 가장 VAR-meanpos 이소 모델에 의해 설명되는 형광. 비슷한 상황에있는 염료는 모델이 적용되는 VAR-meanpos을 자유롭게 회전 할 수 없습니다. 더 라 이 모델에 대한 etails는 우리의 최근 출판물 (32)에서 찾을 수 있습니다.
이 모델은 특히 염료 발생할 수있는 다양한 환경을 설명하고이를 적용하는 것은 현명 현지화 정밀도를 최적화에 광범위한 레퍼토리를 제공합니다. 고속 NPS의 특정 위치에 부착 된 각 염료 분자는 FRET-파트너가 서로 다른 모델이 허용되도록, 각각의 모델에 할당 될 수있다. 이 무한한 및 확대에 자연을 모델링 할 수 있습니다. 그러나, 하나의 최종 모델의 조합에 의해 얻어진 결과는 실험 데이터와 일치 여전히되도록 엄격한 통계 학적 테스트를 수행하는 것이 중요하다. 이 시험은 빨리 NPS 소프트웨어에 포함되어 있습니다.
실험 데이터에 고속 NPS를 적용하기 위해 (단지) 세 개의 입력 변수의 측정이 필요하다. 우선, 색소 쌍 특정 등방성 포스터 반경 (/54782/54782eq5.jpg "/>)를 정의 할 필요가 도너 염료. 따라서, 양자 수율 (QY) 상기 도너 형광 발광 스펙트럼과 수용자의 흡수 스펙트럼을 측정 할 필요가있다. 이러한 측정은 수행 될 수있다 표준 분광기 및 표준 형광 분광기를 사용하여 대량. 각각의 쌍의 경우, R 0은 프리웨어 PhotochemCAD을 사용하여 계산되고, NPS 분석에 사용될 수있다. 또한, 염료 분자 (시간 – 분해) 형광 이방성 필요 편광 (시간)에 민감한 형광 분광기를 사용하여 얻을 수있다. 그러나, 고속 NPS 가장 중요한 입력 파라미터는 전반사 형광 현미경 (TIRFM)와 같은 단일 분자 형광 현미경 설정에서 측정 smFRET 효율이다 .
여기서는 smFRET 데이터를 획득하고 고속 NPS (도 1)를 적용하는 단계별 프로토콜을 제시한다.
