Determinare la struttura di una biomolecola è un prerequisito chiave per comprendere la sua funzione. Due metodi consolidati per la determinazione della struttura sono la microscopia crio-elettroni e raggi X cristallografia 1, 2. Oggi, entrambi i metodi forniscono alta risoluzione informazioni strutturali con risoluzione fino al livello angstrom. Questi due metodi sono stati ampiamente utilizzati per chiarire la struttura di grandi biomolecole come complessi proteici. Anche se i metodi esistenti sono costantemente state migliorate nel corso degli ultimi decenni, la complessità delle strutture biologiche pone ancora una sfida importante per la biologia strutturale, in particolare quando le grandi, dinamiche e transitori complessi sono indagati 3.
Al fine di studiare le dinamiche di complessi macromolecolari e il rapporto struttura-funzione, in particolare, le metodologie a singola molecola hanno provided informazioni utili 4. Diverse nuove strategie sono state sviluppate fornendo un approccio ortogonale di acquisire informazioni strutturali e dinamiche. Esempi sono ad alta velocità AFM 5, manipolazione meccanica 6, a fluorescenza localizzazione microscopia a 7, così come singola molecola trasferimento di energia per risonanza (smFRET) 8, 9. Da molto presto FRET è stato definito un righello molecolare, a causa della dipendenza distanza sulla scala di lunghezza di biomacromolecole 10.
Uno particolarmente interessante applicazione smFRET è quello di utilizzare le informazioni distanza ottenuta da misurazioni smFRET dedurre informazioni 11, 12, 13, 14, 15 strutturale </sup>, 16, 17, 18, 19, 20, 21, 22, 23. Grazie alla elevata risoluzione temporale del smFRET, la posizione delle parti mobili di una struttura proteica può essere localizzato. Tuttavia, al fine di estrarre informazioni quantitative da dati smFRET importanti parametri di correzione sulle molecole di colorante necessario determinare durante la misura 24. Con questi fattori di correzione, l'efficienza FRET E FRET può essere calcolato con la formula
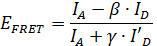 ,
,
dove A e D </sub> sono le intensità di fluorescenza del donatore e la molecola accettore, rispettivamente (vedere Figura 2). I conti β-fattore per il cross-talk, la fuoriuscita di emissione del donatore nel canale accettore ed è calcolato

dove 'A e I' d sono le intensità di fluorescenza del donatore e la molecola accettore dopo fotobleaching della molecola accettore.
La γ-fattore corregge la differenza in termini di efficienza di rilevazione relativi nei due canali nonché le differenze nel rendimento di fluorescenza quantica del donatore e il colorante accettore. Si è calcolata da ogni individuo traccia temporale dai
<p class="jove_content" fo:keep-together.within-page="1" fo:text-align="center" style = "text-align: center;">
Si noti, che questa descrizione trascura eccitazione diretta della molecola accettore, che talvolta diventa importante e avrebbe bisogno di essere corretto per pure. Per determinare tali fattori di correzione è utile per eccitare sia il donatore e l'accettore in uno schema alternato 25 per distinguere tra i cambiamenti foto-fisiche e dinamiche strutturali.
Al fine di ottenere non solo l'efficienza smFRET quantitativi, ma anche informazioni strutturali quantitative, il sistema Nano-posizionamento (NPS) è stato introdotto nel 2008 26. Il nome è stato scelto sulla base delle sue somiglianze con il Global Positioning System basato su satellite (GPS). La NPS è una tecnica ibrida che combina smFRET e X-ray dati cristallografia per la localizzazione di posizioni colorante sconosciuti in complessi biomacromolecular. la cStruttura rystal serve come un quadro di riferimento ei risultati smFRET vengono utilizzati per ottenere informazioni distanza tra una posizione sconosciuta fluoroforo (antenna) e una posizione nota dalla struttura cristallina (satellite). In esperimenti consecutivi le distanze tra l'antenna e diversi satelliti sono misurati e la posizione dell'antenna è determinata mediante un sistema di analisi statisticamente rigoroso basato sulla stima dei parametri bayesiana. Come risultato, non solo la posizione più probabile dell'antenna viene calcolata, ma la sua distribuzione completa incertezza 3D, la cosiddetta posteriori, visualizzata da volumi credibili. Inoltre, NPS è stata ampliata per consentire l'analisi delle reti complete smFRET 27.
La NPS è stato utilizzato per risolvere un certo numero di importanti questioni trascrizione eucariotico, cioè il corso del DNA monte, il DNA non-modello e l'mRNA nascente all'interno della RNA polimerasi II allungamento complex 12, 28, dimostra altresì l'effetto di inizio della trascrizione fattori 26 e l'architettura dinamica di un open-promotore complesso 29. Inoltre, il NPS è stato utilizzato per chiarire la struttura del archaeal RNA polimerasi complesso aperto 30 e in particolare la posizione del fattore di trascrizione iniziazione TFE, che si lega competitivamente allo stesso sito come fattore di trascrizione allungamento Spt4 / 5 31.
Da allora, un certo numero di approcci strutturali basate smFRET state pubblicate 15, 18, 21, 23. Quando si confrontano diversi metodi strutturali basati smFRET, diventa chiaro che l'apparente precisione del metodo è fortemente dipendente dalla particolare scelta di modelli di tintura. Si deve notare chemolecole di colorante possono presentare comportamento spaziale e orientativo diverso a seconda del loro ambiente locale.
A tal fine, fast-NPS stato introdotto 32. Fast-NPS utilizza un algoritmo di campionamento advanced riducendo i tempi di calcolo drasticamente. Inoltre, fast-NPS permette di eseguire una analisi strutturale e per ogni molecola di colorante l'utente può scegliere tra una serie di cinque diversi modelli di tintura che verrà descritto in seguito. Il modello più conservativo, chiamato classico, si assume che il colorante occupa solo una, ma sconosciuto, posizione. In questa posizione, il fluoroforo può ruotare liberamente all'interno di un cono, la cui dimensione è determinata dalla rispettiva (dipendente dal tempo) anisotropia di fluorescenza. L'orientamento del cono non è noto, il che porta a grandi incertezze nella conversione efficienze smFRET misurate in distanze. A questo proposito, il modello è conservatore, poiché porterà alla più piccola precisione rispetto alle altre modalità dyeLS. Solo per distanze molto brevi qualora le ipotesi formulate dal piombo modello classico ad una determinazione della posizione notevolmente errato. Per valori tipici smFRET, la posizione corretta è sempre racchiuso nel comparativamente grande volume credibile.
Tuttavia, poiché una maggiore precisione è desiderabile, è importante sviluppare e testare modelli coloranti alternativi, che potrebbe contribuire a migliorare la precisione. Se il colorante gira molto più velocemente rispetto alla durata della fluorescenza intrinseca, il cosiddetto modello iso può essere applicata. Qui, il fattore di orientamento kappa 2 (necessario per il calcolo del raggio Förster caratteristica isotropo  ) È impostato su 2/3. Come risultato, i volumi credibili calcolate sono quasi due ordini di grandezza inferiore rispetto a quelle del modello classico 32. Nel caso in cui il fluoroforo si trova in un ambiente che consente non solo reori velocesentazione, ma il movimento in più veloce in tutto il suo volume accessibile, il modello meanpos-iso devono essere utilizzati. In questo modello, il colorante occupa effettivamente solo una posizione media, dove la media spaziale è rappresentato da una conversione distanza polinomio 15. Questo modello si applica se per esempio il colorante (comunemente idrofobica) è collegata ad una regione idrofila, per esempio, il DNA. Applicazione del modello meanpos-iso comporta un'ulteriore riduzione delle dimensioni dei volumi credibili per un fattore di circa due. Tuttavia, un colorante legata ad una proteina potrebbe legarsi reversibilmente a diverse patch idrofobiche suo volume stericamente accessibile (AV). Un fluoroforo che passa istantaneamente tra queste regioni, ma all'interno di una regione sottoposta a rotazione libera e movimento veloce localizzata è meglio descritta dal modello VAR-meanpos-iso. Per una situazione simile in cui il colorante non è libero di ruotare i VAR-meanpos modello si applica. più d etails su questi modelli si possono trovare nella nostra recente pubblicazione 32.
) È impostato su 2/3. Come risultato, i volumi credibili calcolate sono quasi due ordini di grandezza inferiore rispetto a quelle del modello classico 32. Nel caso in cui il fluoroforo si trova in un ambiente che consente non solo reori velocesentazione, ma il movimento in più veloce in tutto il suo volume accessibile, il modello meanpos-iso devono essere utilizzati. In questo modello, il colorante occupa effettivamente solo una posizione media, dove la media spaziale è rappresentato da una conversione distanza polinomio 15. Questo modello si applica se per esempio il colorante (comunemente idrofobica) è collegata ad una regione idrofila, per esempio, il DNA. Applicazione del modello meanpos-iso comporta un'ulteriore riduzione delle dimensioni dei volumi credibili per un fattore di circa due. Tuttavia, un colorante legata ad una proteina potrebbe legarsi reversibilmente a diverse patch idrofobiche suo volume stericamente accessibile (AV). Un fluoroforo che passa istantaneamente tra queste regioni, ma all'interno di una regione sottoposta a rotazione libera e movimento veloce localizzata è meglio descritta dal modello VAR-meanpos-iso. Per una situazione simile in cui il colorante non è libero di ruotare i VAR-meanpos modello si applica. più d etails su questi modelli si possono trovare nella nostra recente pubblicazione 32.
Questi modelli offrono un vasto repertorio di conto specifico per i vari ambienti di un colorante potrebbero incontrare e loro applicazione ottimizza saggiamente la sua precisione di localizzazione. In Fast-NPS ogni molecola di colorante attaccato ad una posizione specifica può essere assegnato a un singolo modello, in modo tale che FRET-partner sono autorizzati ad avere modelli diversi. Questo consente la modellazione illimitata e vicino alla natura. Tuttavia, è importante che si compie test statistici rigorosi per garantire che il risultato ottenuto dalla combinazione modello finale è ancora in accordo con i dati sperimentali. Questi test sono inclusi nel software Fast-NPS.
Per applicare Fast-NPS ai dati sperimentali è richiesta la misurazione (solo) tre parametri di input. In primo luogo, il colorante-coppia specifica isotropo Förster raggi (/54782/54782eq5.jpg "/>) Devono essere determinate. Pertanto, la resa quantica (QY) del colorante donatori, gli spettri di emissione di fluorescenza del donatore e lo spettro di assorbimento accettore devono essere misurate. Queste misurazioni possono essere effettuate in bulk, utilizzando uno spettrometro standard ed uno spettrometro a fluorescenza. Per ogni coppia, l'R 0 viene quindi calcolato utilizzando la PhotochemCAD freeware e può essere usato nell'analisi NPS. Inoltre, il (tempo risolto) anisotropie fluorescenza delle molecole di colorante bisogno essere ottenuti utilizzando una polarizzazione (e tempo) spettrometro di fluorescenza sensibile. Tuttavia, i parametri di input più importanti per fast-NPS sono le efficienze smFRET misurati su una configurazione microscopia a fluorescenza singola molecola, come un microscopio riflessione a fluorescenza interna totale (TIRFM) .
Qui, vi presentiamo un protocollo step-by-step per ottenere dati smFRET e l'applicazione di Fast-NPS (Figura 1).
