La détermination de la structure d'une biomolécule est une condition essentielle pour la compréhension de sa fonction. Deux méthodes bien établies pour la détermination de la structure sont cryo-microscopie électronique et cristallographie aux rayons X 1, 2. Aujourd'hui, les deux méthodes fournissent à haute résolution des informations structurelles avec une résolution jusqu'au niveau angström. Ces deux méthodes ont été largement utilisées pour élucider la structure de grandes biomolécules telles que des complexes protéiques. Bien que les méthodes existantes ont été constamment améliorées au cours des dernières décennies, la complexité des structures biologiques pose toujours un défi majeur pour la biologie structurale, en particulier lorsque de grandes, dynamiques et transitoires complexes sont étudiés 3.
Afin d'étudier la dynamique des complexes macromoléculaires et la relation structure-fonction, en particulier, les méthodologies d'une seule molécule ont provided informations utiles 4. Plusieurs nouvelles stratégies ont été élaborées fournissant une approche orthogonale sur l'acquisition de l'information structurelle et dynamique. Les exemples sont AFM à grande vitesse 5, manipulation mécanique 6, fluorescence localisation microscopie 7, ainsi que une seule molécule de transfert Förster Resonance Energy (smFRET) 8, 9. Depuis très tôt FRET a été appelé une règle moléculaire, en raison de la dépendance de la distance sur l'échelle de longueur de biomacromolecules 10.
Une application particulièrement intéressante de smFRET consiste à utiliser l'information de distance obtenue à partir des mesures smFRET pour déduire des informations de structure 11, 12, 13, 14, 15 </sup>, 16, 17, 18, 19, 20, 21, 22, 23. En raison de la haute résolution temporelle de smFRET, la position des parties mobiles d'une structure protéique peut être localisée. Toutefois, afin d' en extraire des informations quantitatives de données smFRET correction des paramètres importants sur les molécules de colorant doivent être déterminés lors de la mesure 24. Avec ces facteurs de correction, l'efficacité de FRET E FRET peut être calculée en utilisant la formule
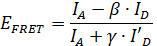 ,
,
où je A et I D </sub> sont les intensités de fluorescence du donneur et la molécule accepteur, respectivement (voir la figure 2). Comptes Le β-facteur de la diaphonie, la fuite d'émission du donneur dans le canal accepteur et est calculée par

où I 'A et je' D sont les intensités de fluorescence du donneur et la molécule accepteur après photo blanchiment de la molécule d'accepteur.
Γ le facteur corrige la différence dans les efficacités de détection relatif dans les deux canaux, ainsi que les différences dans le rendement quantique de fluorescence du donneur et le colorant accepteur. Il est calculé à partir de toute trace de temps individuel par
<p class="jove_content" fo:keep-together.within-page="1" fo:text-align="center" style = "text-align: center;">
Notez que cette description néglige excitation directe de la molécule accepteur, qui devient parfois importante et aurait besoin d'être corrigée pour ainsi. Pour la détermination de ces facteurs de correction , il est utile d'exciter le donneur ainsi que l'accepteur dans un schéma alternatif 25 afin de différencier entre les changements de photo-physique et dynamique des structures.
Afin d'obtenir non seulement l' efficacité de smFRET quantitatives , mais aussi des informations structurelles quantitative, le système Nano-positionnement (NPS) a été introduit en 2008 26. Le nom a été choisi en fonction de ses similitudes avec le système de positionnement global par satellite (GPS). Le NPS est une technique hybride combinant smFRET et les données de cristallographie aux rayons X pour la localisation des positions de colorants inconnus dans les complexes biomacromolecular. Le cStructure rystal sert de cadre de référence et les résultats smFRET sont utilisés pour obtenir des informations de distance entre une position inconnue fluorophore (antenne) et une position connue à partir de la structure cristalline (satellite). Dans des expériences successives les distances entre l'antenne et plusieurs satellites sont mesurés et la position de l'antenne est déterminée au moyen d'une méthode d'analyse statistique rigoureuse basée sur Bayesian estimation des paramètres. Par conséquent, non seulement la position la plus probable de l'antenne est calculé, mais la distribution de l'incertitude 3D complet, la partie postérieure dite, visualisée par des volumes crédibles. En outre, NPS a été élargi pour permettre l'analyse des réseaux complets de smFRET 27.
Le NPS a été utilisé pour résoudre un certain nombre de questions importantes dans la transcription eucaryote, à savoir le cours de l'ADN en amont, l'ADN non-modèle et l'ARNm naissant au sein de l'allongement co RNA Polymerase IImplex 12, 28, ce qui démontre également l'effet de la transcription facteurs d' initiation 26 et l'architecture dynamique d'un open-promotor complexe 29. En outre, le NPS a été utilisé pour élucider la structure de l'Archaea complexe ouvert RNA Polymerase 30 et en particulier la position du facteur initiation de la transcription TFE, qui se lie de manière compétitive à le même site que le facteur de transcription de l' allongement Spt4 / 5 31.
Depuis lors, un certain nombre d'approches structurelles fondées smFRET ont été publiés 15, 18, 21, 23. Lorsque l'on compare les différentes méthodes structurelles fondées smFRET, il devient clair que la précision apparente de la méthode est fortement dépendante du choix particulier de modèles de colorant. Il faut noter quedes molécules de colorant peuvent présenter un comportement spatial et d'orientation différente en fonction de leur environnement local.
À cette fin, Fast-NPS a été introduit 32. Fast NPS utilise un algorithme d'échantillonnage avancé réduisant les temps de calcul de manière drastique. En outre, Fast-NPS permet d'effectuer une analyse structurelle et pour chaque molécule de colorant l'utilisateur peut choisir parmi un ensemble de cinq modèles de colorants différents qui sera décrit suivant. Le modèle le plus conservateur, appelé classique, suppose que le colorant occupe une seule, mais inconnue, la position. Dans cette position, le fluorophore peut tourner librement à l'intérieur d'un cône dont la taille est déterminée à partir de son respectif (dépendant du temps) anisotropie de fluorescence. L'orientation du cône est inconnue, ce qui conduit à des incertitudes importantes lors de la conversion de l'efficacité smFRET mesurées en distances. À cet égard, le modèle est conservatrice, car elle conduira à la plus petite précision par rapport à l'autre mode de colorantls. Seulement pour les très courtes distances devrait les hypothèses retenues par le modèle classique de plomb à une détermination de la position sensiblement incorrecte. Pour les valeurs typiques de smFRET, la position correcte est toujours enfermé dans le volume relativement important crédible.
Cependant, comme une plus grande précision est souhaitable, il est important de développer et de tester des modèles de colorants alternatifs, qui pourraient aider à améliorer la précision. Si le colorant tourne beaucoup plus rapidement que sa durée de vie de fluorescence intrinsèque, le soi-disant modèle iso peut être appliqué. Ici, le facteur d'orientation k 2 (nécessaire pour le calcul du rayon de Förster isotrope caractéristique  ) Est fixé à 2/3. Par conséquent, les volumes calculés sont fiables presque deux ordres de grandeur par rapport à celles du modèle classique 32. Dans le cas où le fluorophore se trouve dans un environnement qui permet non seulement rapide reorientation, mais le mouvement plus rapide sur tout son volume accessible, le modèle meanpos-iso devraient être utilisés. Dans ce modèle, le colorant de manière efficace que occupe une position moyenne, où la moyenne spatiale est représentée par une conversion de 15 minutes polynomiale. Ce modèle est applicable par exemple si le colorant (généralement hydrophobe) est fixé à une région hydrophile, par exemple, l'ADN. Application du modèle meanpos-iso conduit à une réduction supplémentaire de la taille des volumes crédibles par un facteur d'environ deux. Cependant, un colorant lié à une protéine peut se lier de manière réversible à plusieurs taches hydrophobes dans son volume stériquement accessibles (AV). Un fluorophore qui bascule instantanément entre ces régions, mais dans une région subit une rotation libre et le mouvement localisé rapide est mieux décrit par le modèle var-meanpos-iso. Pour une situation similaire dans laquelle le colorant est pas libre de faire pivoter les var-meanpos modèle applique. Plus d ÉTAILS sur ces modèles peuvent être trouvés dans notre récente publication 32.
) Est fixé à 2/3. Par conséquent, les volumes calculés sont fiables presque deux ordres de grandeur par rapport à celles du modèle classique 32. Dans le cas où le fluorophore se trouve dans un environnement qui permet non seulement rapide reorientation, mais le mouvement plus rapide sur tout son volume accessible, le modèle meanpos-iso devraient être utilisés. Dans ce modèle, le colorant de manière efficace que occupe une position moyenne, où la moyenne spatiale est représentée par une conversion de 15 minutes polynomiale. Ce modèle est applicable par exemple si le colorant (généralement hydrophobe) est fixé à une région hydrophile, par exemple, l'ADN. Application du modèle meanpos-iso conduit à une réduction supplémentaire de la taille des volumes crédibles par un facteur d'environ deux. Cependant, un colorant lié à une protéine peut se lier de manière réversible à plusieurs taches hydrophobes dans son volume stériquement accessibles (AV). Un fluorophore qui bascule instantanément entre ces régions, mais dans une région subit une rotation libre et le mouvement localisé rapide est mieux décrit par le modèle var-meanpos-iso. Pour une situation similaire dans laquelle le colorant est pas libre de faire pivoter les var-meanpos modèle applique. Plus d ÉTAILS sur ces modèles peuvent être trouvés dans notre récente publication 32.
Ces modèles offrent un vaste répertoire pour tenir compte spécifiquement pour les différents milieux un colorant pourrait rencontrer et de les appliquer à bon escient optimise la précision de la localisation. Dans rapide NPS chaque molécule de colorant attaché à une position spécifique peut être affecté à un modèle individuel, de telle sorte que FRET-partenaires sont autorisés à avoir des modèles différents. Cela permet la modélisation sans limites et à proximité à la nature. Cependant, il est important que l'on effectue des tests statistiques rigoureux pour assurer que le résultat obtenu par la combinaison du modèle final est toujours en accord avec les données expérimentales. Ces tests sont inclus dans le logiciel rapide NPS.
Pour appliquer Fast NPS aux données expérimentales de la mesure de (seulement) trois paramètres d'entrée est nécessaire. Tout d'abord, le colorant paire isotrope Förster spécifique rayons (/54782/54782eq5.jpg "/>) Doivent être déterminés. Par conséquent, le rendement quantique (QY) du colorant donneur, les spectres d'émission de fluorescence du donneur et le spectre d'absorption de l'accepteur doivent être mesurés. Ces mesures peuvent être réalisées en en masse, en utilisant un spectromètre standard et d' un spectromètre à fluorescence standard. Pour chaque couple, le R 0 est ensuite calculée en utilisant l'PhotochemCAD gratuit et peut être utilisé dans l'analyse SNP. Par ailleurs, les (résolu temps) anisotropies de fluorescence des molécules de colorant doivent être obtenue en utilisant une polarisation (et le temps), un spectromètre de fluorescence sensible. Toutefois, les paramètres d'entrée les plus importants pour fast NPS sont les rendements smFRET mesurés sur une installation de microscopie à fluorescence de molécule unique, comme une interne fluorescence par réflexion microscope totale (TIRFM) .
Ici, nous présentons un protocole étape par étape pour obtenir des données smFRET et l' application rapide NPS (Figure 1).
