Five male C57BL/6J were implanted with an ECoG headset and headplate (Figure 4A). After recovery, mice were habituated to head-fixation and the electrophysiology recording rig during two 1.5 h sessions on separate days (Figure 4B). Next, a 2 mm x2 mm craniotomy window was created (Figure 4C) and a silicon probe was inserted with the mouse awake and head-fixed (Figure 4D). Two types of silicon UCLA probes were used: (1) 64 M: a one-shank, 64 electrode probe; and (2) 256 ANS: a four-shank, 256 electrode probe. Sevoflurane and oxygen were administered using a nose cone fixed to the electrophysiology rig. Levels of oxygen and sevoflurane were monitored every 5 min using an inline gas analyzer. Following transcardial perfusion, the brain was harvested (Figure 4E) and evaluated for probe trajectory (Figure 4F). Recorded action potentials were then curated into single unit data (Figure 4G).
Five recordings were performed wherein probe placement was validated with available probe reconstruction algorithms developed in MATLAB23. Using these algorithms, each electrode of the silicon probe was localized to a specific brain structure so that a three dimensional perspective of probe placement within the brain was acquired (Figure 5). These reconstruction algorithms were coupled with immunohistochemical markers to further validate probe trajectories (Figure 6). The data was then matched with curated single units (Figure 7).
A total of 64 single neurons was captured, of which 13 were located in the vlPAG. The remaining 51 neurons were in nearby nuclei: midbrain, midbrain reticular nucleus, paratrochlear nucleus, cuneiform nucleus, laterodorsdal tegmental nucleus, parabrachial nucleus, and superior cerebellar peduncles. The firing activity of neurons in each mouse is presented as heat maps (Figure 7). Mouse 3 was excluded from the analysis because of poor quality single-unit recording. Most recorded neurons decreased their firing during sevoflurane anesthesia.
Mean firing of vlPAG neurons was compared for each mouse at 5 min baseline (i.e., 10-15 min into the recording) and 5 min during sevoflurane (i.e., 40-45 min into the recording). vlPAG firing was significantly decreased (Figure 8A) and furthermore, this increase was consistent across all vlPAG neurons (Figure 8B).
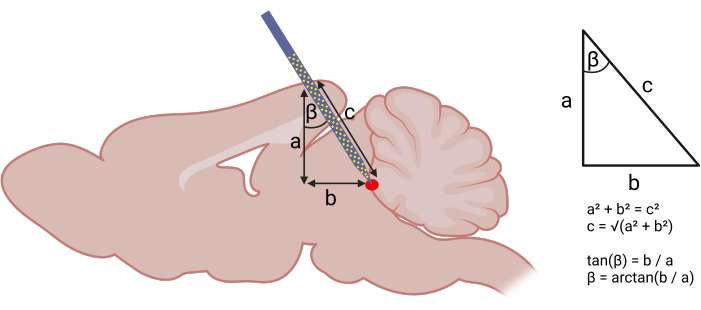
Figure 1: Calculating stereotaxic coordinates for probe insertion. Left panel: Using geometry to calculate angled approach for probe insertion. The following components of a right triangle were identified: (a) DV depth of the target structure from the brain surface (acquired using the stereotaxic atlas), (b) distance between the AP coordinate assuming a strictly vertical descent and the AP coordinate of the target structure. Right panel: With (a) and (b), one can use the Pythagorean theorem to calculate the AP angle (β) and length (c) of probe insertion. The following coordinates were used to target the vlPAG: (AP) -3.6 mm, (ML) +0.5 mm, (DV) -4 mm. An AP angle (β) of 20° was used. Please click here to view a larger version of this figure.

Figure 2: Headplate implantation and single unit recording. (A) ECoG headset wire placement during implantation. (B) Top view of the mouse skull after ECoG headset and headplate implantation. (C) Silicon probe shanks inserted into the brain.(D) Head-fixed mouse during a silicon probe recording. Please click here to view a larger version of this figure.
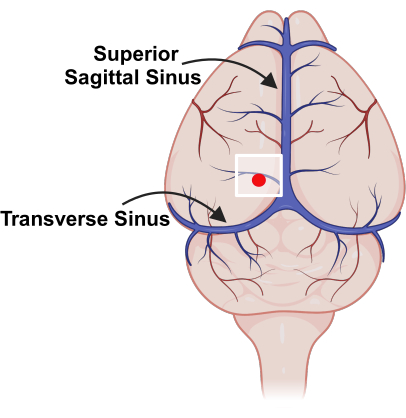
Figure 3: Placement of cranial window, relative to major blood vessels. Schematic representation of vasculature on the surface of the mouse brain with superior sagittal sinus and transverse sinus marked. The white square indicates the location of the cranial window and the red dot represents the probe insertion point. Please click here to view a larger version of this figure.
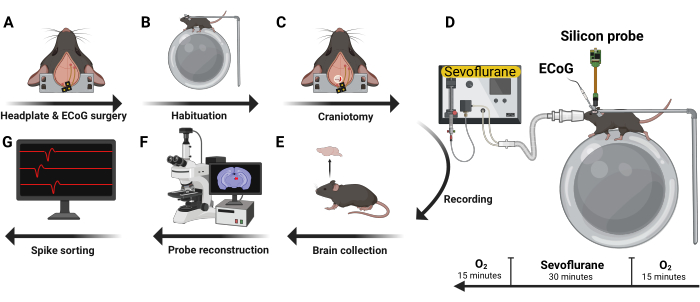
Figure 4: Experimental design. Five male C57BL/6J mice were implanted with ECoG headset and headplate (A). After recovery, they were habituated to head-fixation and the recording rig (B). Next, a craniotomy was performed (C) and a silicon probe inserted with the mouse awake and in a head-fixed position. Volatile anesthetics and oxygen were administered using a nose cone (D). After recording, the brain was harvested (E) and analyzed using immunohistochemistry techniques to reconstruct the trajectory of the probe (F). Recorded action potentials were then curated into single unit data (G). Please click here to view a larger version of this figure.
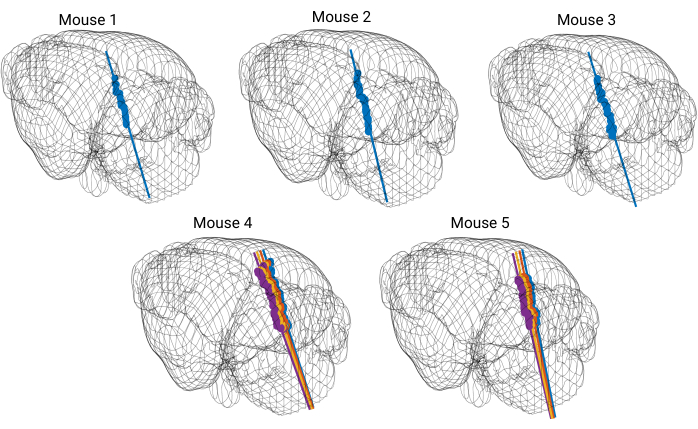
Figure 5: 3D model of the brain with reconstructed probe tracks. Each straight colored line represents a probe trajectory. Each dot marks the location of DiI dye visible on coronal brain slices. The probe trajectory is reconstructed for every shank of the silicon probe. Please click here to view a larger version of this figure.
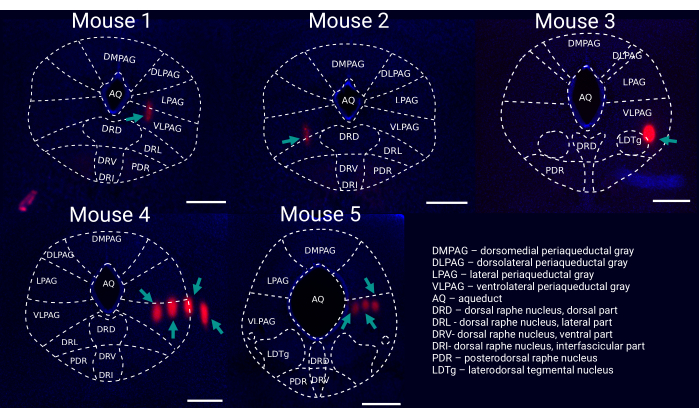
Figure 6: Post hoc reconstruction of probe placement within the vlPAG. Representative coronal brain slice for each recorded mouse (1-5) stained with DAPI (blue) showing probe tracks ((DiI, red, marked by green arrows) with an outline of brain regions. Scale bar: 500 µm. Mouse 1-3 were recorded with single shank silicon probes (i.e., 64 channels), whereas mouse 4 & 5 were recorded with four-shank silicon probes (i.e., 256 channels). Please click here to view a larger version of this figure.
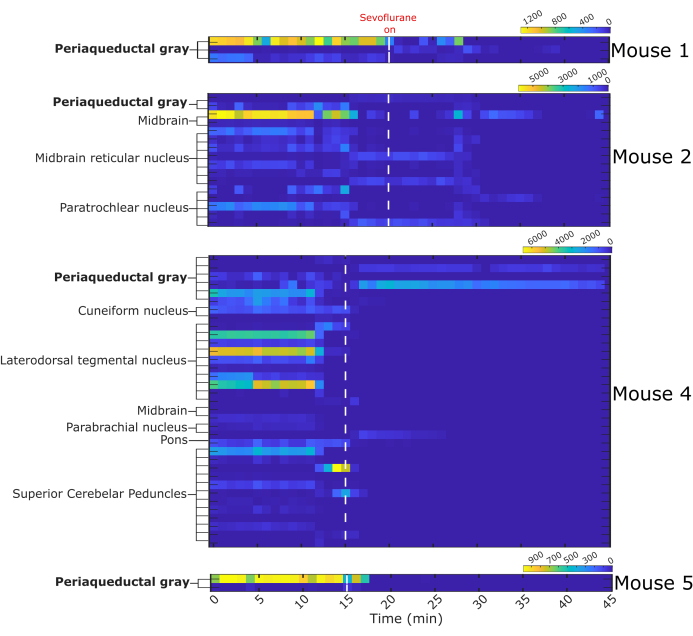
Figure 7: Binned firing activity before and during sevoflurane anesthesia. Heat maps represent the number of fired action potentials by 64 single neurons in one-minute intervals. Single neurons were assigned a structure of origin based on the probe trajectory reconstruction. White dashed line indicates start of sevoflurane administration, n = 4 mice. Please click here to view a larger version of this figure.
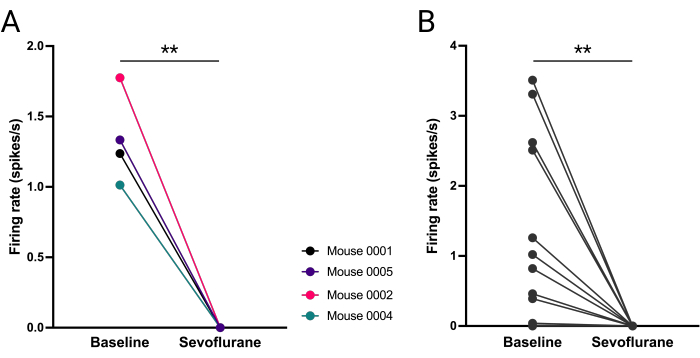
Figure 8: Abolished firing of vlPAG neurons during sevoflurane anesthesia. (A) Mean number of action potentials fired before and during anesthesia for each mouse. (B) Number of action potentials fired by each vlPAG neuron (B). Paired t-student test, **P = 0.0036, P = 0.0053 respectively. Please click here to view a larger version of this figure.
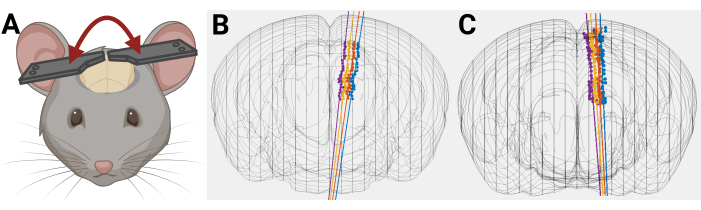
Figure 9: Example of how uneven headplate placement can affect probe trajectory. (A) Uneven placement of headplate on mouse head. (B,C) 3D mouse brain reconstruction with rendered probe trajectory where there is an unintentional mediolateral angle caused by uneven placement of headplates on mouse head. Please click here to view a larger version of this figure.
