Figure 4 shows a representative example of the results obtained when protocol 1 was applied to a single subject (46 years old, female). PI was increased soon after having chewed (T7) both a hard (from 1.73 numb/s to 2.27 numb/s) and soft pellet (from 1.67 numb/s to 1.87 numb/s) (Figure 4A). However, 30 min later (T37), the increased performance persisted only for the hard pellet. On the other hand, both a lack of activity and the handgrip exercise had a negative effect on performance, which dropped from 1.73 numb/s to 1.67 numb/s and from 1.6 numb/s to 1.53 numb/s, with a tendency to recover observed 30 min later, during the last experimental evaluation.
As observed in Figure 4B, qualitatively similar changes were observed for the task-related mydriasis. In this instance, measurements consisted of individual samples taken randomly when the subject was resting. During the haptic task, two samples were recorded, but the first was discarded. Alternatively, in the continuous acquisition mode of the instrument, 100 samples were recorded in 20 s, with the first 20–50 measurements disregarded, and the remaining were then averaged following the removal of blink artefacts (Figure 2C,D). Individual samples closely reflect the average value, due to the fact that pupil size reaches a very stable level 4–10 s following the switching off of eye illumination (Figure 2C,D). Data illustrated in Figure 4 and Figure 5 have been replicated in a population of 30 subjects, and both the chewing- and handgrip-induced changes were statistically confirmed. On the other hand, when the subjects were not involved in any activity, there were no modifications in cognitive performance and mydriasis30 both at T7 and T37.
Despite the fact that 1) performance and mydriasis were recorded in different tasks and 2) the 12 experimental points illustrated in Figure 5A,B were recorded on 4 separate days, it is remarkable that a strong correlation was observed between performance and task-related mydriasis (r = 0.939, p < 0.0005, y = 1.166x – 0.417). As can be inferred from Figure 5A, this relation was due to the modifications induced by chewing hard and soft pellets. Even more surprisingly, a correlation was evident also when the corresponding changes with respect to baseline values were considered (r = 0.924, p < 0.001, y = 1.210x + 0.101; Figure 5B).
Among the 30 subjects analyzed in the study of Tramonti Fantozzi et al.30, PI and mydriasis were significantly correlated in 26 of them, with slopes of the corresponding regression lines ranging from 0.310–1.327 numb/s/mm. The corresponding changes were significantly correlated in 22 subjects (range of slopes: 0.390–1.408).
Even stronger evidence of LC involvement in the stimulating effects of chewing on cognitive performance can be obtained by correlating the chewing-induced changes in PI with the change in mydriasis observed only during the execution of the matrices test. This can be achieved under the more natural conditions of protocol 2, in which subjects perform the matrices test while pupil size is simultaneously recorded (Figure 6).

Figure 1: Example of Spinnler-Tognoni numerical matrices. The test consists in identifying the target numbers indicated above each matrix, which have been ticked by the subject. Please click here to view a larger version of this figure.
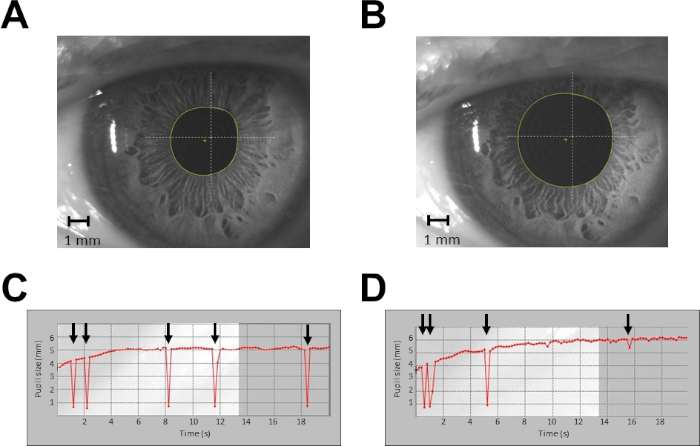
Figure 2: Example of pupil size recordings from a single subject in protocol 1. (A) Recording of pupil size at rest, single shot. (B) Recording of pupil size during haptic task, single shot. (C) Continuous recording of pupil size at rest for 20 s. (D) Continuous recording of pupil size during haptic task for 20 s. Arrows indicate blinking artefacts. In (C) and (D), data taken from time 0 to time 4 s are discarded from the analysis. Please click here to view a larger version of this figure.
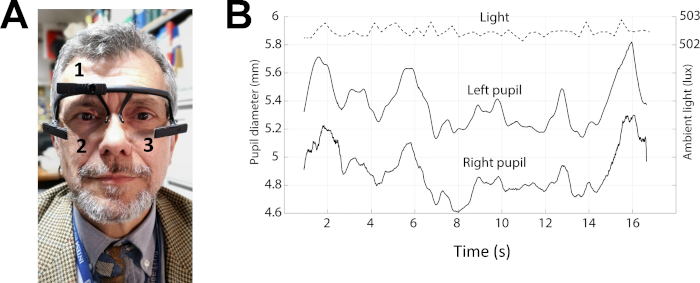
Figure 3: Example of pupil size recordings in protocol 2. (A) Photo of a subject wearing the pupillometer. The numbers 1–3 indicates the position of the three cameras, which allow behaviour (1) and pupil size (2-3) recordings. (B) Top trace: level of the environmental lightening. Middle and bottom traces: left and right pupil size during performance of the Spinnler-Tognoni matrices test. Please click here to view a larger version of this figure.
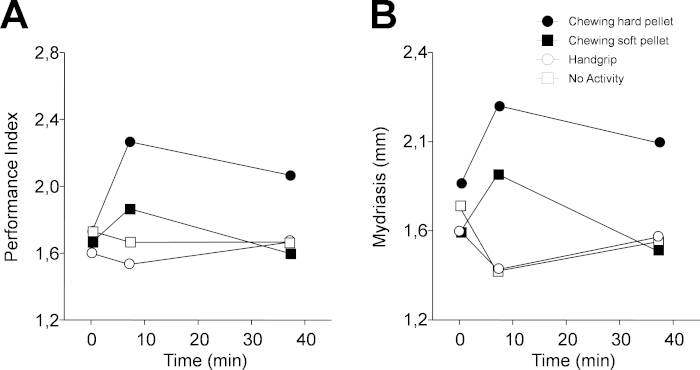
Figure 4: Changes in performance and task-related mydriasis induced by different sensorimotor activities in protocol 1. (A) Changes in PI. (B) Changes in task-related mydriasis. In (A) and (B), dots, black squares, circles and white squares represent data relative to chewing hard pellet, chewing soft pellet, handgrip, and no activity, respectively. Each activity was performed for 2 min from time 5 min to time 7 min. Please click here to view a larger version of this figure.
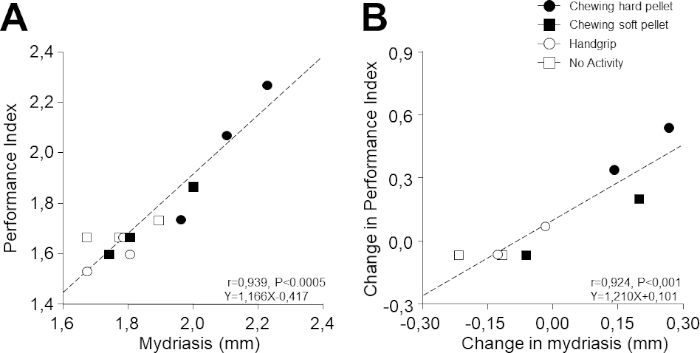
Figure 5: Relation between PI and task-related mydriasis. (A) PI values obtained at different times during the different activities illustrated in Figure 4 are plotted as a function of the corresponding values of task-related mydriasis. (B) Changes in PI with respect to T0 (evaluated as a difference) have been plotted as a function of the corresponding changes in task-related mydriasis. In (A) and (B), dots, black squares, circles, and white squares represent data relative to chewing hard pellet, chewing soft pellet, handgrip, and no activity, respectively. Dashed lines are regression lines of all the data points. Please click here to view a larger version of this figure.

Figure 6: Simultaneous recording of performance and task-related mydriasis. Single frame view of a subject performing the attentive matrices test, taken from the camera mounted on the pupilometer frame. The inset on the right upper corner shows the simultaneous images of both pupils. The green circle represents the fixation point. The red spot and circles drown on the pupil are the pupil centre and contour, as evaluated by the tracking system operating on the eye's videos. Please click here to view a larger version of this figure.
