The present study was performed in compliance with the UCLA Institutional Review Board (IRB) protocols and approved by the UCLA Human Subjects Protection Committee. All participants gave written informed consent in order to enroll in this study.
1. Participant Selection
- Obtain IRB approval to perform the study.
- Screen individuals aged 55 and older for cognitive decline using a standardized neuropsychological battery. Include tests of General Intelligence (Subtests of the WAIS-III)13, Fluency (Fruits and Vegetables)14, Attention (Digits Forward and Backward)13, Language (Boston Naming Test)15, Verbal Memory (Buschke-Fuld Selective Reminding Task)16, WMS-III Logical Memory and Verbal Paired Associates learning13, and Visual Memory (Rey-Osterrieth Figure test)17.
- Have the participants complete mood questionnaires such as the Hamilton Depression and Anxiety Inventories18,19 as well as the Mini Mental State Exam (MMSE)20.
- Include participants that score 26 or above on the MMSE and perform better than two standard deviations below normal for their age on cognitive tests. Exclude participants with clinical anxiety, depression or any other neuropsychiatric or neurological illness. Exclude participants who do not meet MRI safety criteria or who do not consent to a blood draw.
NOTE: In the present study, 93 participants met these criteria (mean age = 67.4 years, 31M/49F).
2. Genotyping
- Have a trained phlebotomist or other medical professional draw blood from each participant.
- Isolate 200 µg genomic DNA from 10 mL of the sample as described21.
- Carry out single nucleotide polymorphism (SNP) genotyping using real-time PCR at two loci, rs429358 and rs7412 to discriminate APOE alleles22.
- Incorporate reporter dyes for rs429358 and rs7412 into a SNP genotyping assay. After each PCR amplification cycle is completed, plot fluorescent signals on a graph showing distribution of reporter and quencher dyes. Perform the experiment in duplicate to confirm results.
- Analyze the SNP genotyping data using a software package developed for the real-time PCR procedure output23.
NOTE: The program used in the present study calculates the affinity of the sample to one of the two reporter dyes that, in turn, represents one APOE SNP over the other. In the present study, 34 carriers of the AD risk allele, APOEε4 (heterozygous ε3/ε4) and 46 non-carriers (homozygous ε3/ε3) were enrolled for a total of 80 study participants. Exclude carriers of the APOEε2 allele because there is evidence that this allele may have a protective effect related to AD.
3. Functional and Structural Imaging Data Collection
- Use a 3 Tesla (3T) MRI system to acquire whole-brain imaging data.
- For functional imaging, collect axial slices using an echo planar imaging (EPI) sequence. To facilitate registration of the functional images, acquire axial slices of T2-weighted, co-planar structural images. For high-resolution structural imaging, collect axial slices using a 3D T1-weighted sequence.
NOTE: In the present study, a 3T magnet was used with a 12-channel head coil. The parameters below were designed for a specific scanner and coil. See Table of Materials for more information.- Acquire functional imaging data using the following sequence parameters: repetition time (TR) = 2,500 ms, echo time (TE) = 21 ms, field of view (FOV) = 200 mm x 200 mm, flip angle = 75°, matrix = 64 x 64, 33 slices, slice thickness = 3 mm, interslice gap = 0.75 mm, voxel size = 3.125 x 3.125 x 3.75 mm.
- Trigger the unrelated words associative memory task to begin with the third volume of the functional imaging sequence. To account for steady-state equilibrium, exclude the first two volumes of each functional scan from analyses.
NOTE: The unrelated words associative memory task has been described elsewhere12,24. Briefly, it is a block-design functional task with encoding and retrieval blocks. Participants are instructed to learn pairs of unrelated words. - Acquire T2-weighted, co-planar structural imaging data using the following sequence parameters: TR = 5,000 ms, TE = 34 ms, FOV = 200 mm x 200 mm, flip angle = 90°, matrix = 128 x 128, 28 slices, slice thickness = 3 mm, interslice gap = 1 mm and voxel size = 1.56 x 1.56 x 4 mm.
- Acquire high-resolution structural (anatomical) imaging using the following Magnetization Prepared Rapid Gradient Echo (MPRAGE) sequence parameters: TR = 1,900 ms, TE = 2.26 ms, TI = 900 ms, FOV = 250 mm x 218 mm, flip angle = 9°, matrix = 256 x 215, 176 slices, slice thickness = 1 mm, zero-filled to a matrix of 256 x 224 resulting in a voxel size = 1 x 0.976 x 0.976 mm.
- For functional imaging, collect axial slices using an echo planar imaging (EPI) sequence. To facilitate registration of the functional images, acquire axial slices of T2-weighted, co-planar structural images. For high-resolution structural imaging, collect axial slices using a 3D T1-weighted sequence.
4. fMRI BOLD Data Preprocessing
- Preprocess the functional data using Functional MRI of the Brain (FMRIB) Software Library (FSL) version 6.0 (http://fsl.fmrib.ox.ac.ul) as follows:
- For each participant's dataset, remove head motion artifact from the data using the Motion Correction FMRIB's Linear Image Registration Tool (MCFLIRT)25.
- Remove non-brain tissue from the images using brain extraction tool (BET) with the optional -F flag26.
- Use the FSL Motion Outliers tool to identify any volumes in the functional data where there is excessive motion based on frame displacement between volumes. Flag volumes where motion is measured as an outlier (above the 75th percentile + 1.5 times the inter-quartile range) compared to the rest of the scan and use the output of this program to downweight those volumes in analyses.
NOTE: Before running group comparisons, check that average motion, as measured by FSL Motion Outliers, does not differ across the two groups. This will help ensure that findings are not driven by group-related differences in motion.
- Set up the preprocessing and first-level general linear model (GLM) using the graphical user interface (GUI) for FSL fMRI Expert Analysis Tool (FEAT) for the first participant.
NOTE: Repeat this step for each study participant. To save time, after setting up one run for one participant, write a script to run preprocessing for the remaining study participants' data by altering the "design.fsf" file (FSL FEAT output) for each participant to reference that participant's specific data.- In the data tab, click on "add 4D data" and navigate to the motion-corrected and brain-extracted file. Set the TR to 2.5 s (corresponding to the TR of the functional sequence acquired). Use the default high pass filter (set to 100 s).
NOTE: High pass filtering will remove low frequency signals of no interest. - In the pre-stats tab, click "none" under "motion correction" (as it was already performed in step 4.1). Uncheck "BET brain extraction" (as it was already completed in step 4.1). Type "5" in the box to set 5 mm full-width half-maximum (FWHM) Gaussian kernel for spatial smoothing.
NOTE: The FWHM for the smoothing kernel should generally be set at about twice the size of the functional scan voxel size. - Use the output (6 columns, rows = # of TRs in the scan) of MCFLIRT to create 6 single-column text files that describe the motion correction performed at each volume within the dataset. These will be added to the model as regressors in the next step.
- In the stats tab under "full model setup", add the 6 motion parameters and their temporal derivatives as regressors or explanatory variables (EVs) in the GLM. For each motion EV choose "custom" (1 entry per volume) for basic shape, "none" for convolution and check "apply temporal filtering."
NOTE: Motion parameters do not need to be convolved by any function because they reference the realignment performed at each functional volume during motion correction and thus do not need to be adjusted.
- In the stats tab under "full model setup", add the 6 motion parameters and their temporal derivatives as regressors or explanatory variables (EVs) in the GLM. For each motion EV choose "custom" (1 entry per volume) for basic shape, "none" for convolution and check "apply temporal filtering."
- In the stats tab, select the output of FSL Motion Outliers from step 4.1 under the "add additional confound EVs".
NOTE: This output is a matrix denoting each volume that was flagged for excessive motion and, by adding the confound file, will be deweighted in the GLM. - In the stats tab, click "full model set-up". Create the task timing text files denoting the onset and offset of different task phases and add these as EVs in the GLM by choosing 1 column format and navigating to the relevant text file (include one for the encoding phase of the task and one for the retrieval phase). For "convolution" choose the "double-gamma HRF" option from the drop down list for both of them. Do not model the baseline or non-active portions of the task in the GLM.
NOTE: HRF stands for hemodynamic response function. Convolving the task EV by the HRF shifts the timing of the task EV to be more consistent with expected task-induced BOLD signal changes in the brain. - In the registration tab, check "expanded functional image" and "main structural image" for a two-step registration.
- Select the participant's co-planar T2-weighted structural scan for the first step, in which functional data is registered to the co-planar structural data. Choose 6 degrees of freedom (DOF) for this step by clicking on the second drop down box under this step and choosing "6 DOF".
- For the next step, in which the T2-weighted image is registered to the high resolution T1-weighted MPRAGE, select boundary based registration (BBR) from the drop down box27.
NOTE: BBR uses intensity differences between white matter and gray matter to register structural and functional scans and has been shown to perform better than FLIRT and other alternative methods. - For the final step, in which the high-resolution structural data is registered to the standard MNI152 template, choose 12 degrees of freedom and a linear transformation by choosing "12 DOF".
NOTE: When all the steps in section 4 are complete the functional data are preprocessed and ready for further analysis.
- In the data tab, click on "add 4D data" and navigate to the motion-corrected and brain-extracted file. Set the TR to 2.5 s (corresponding to the TR of the functional sequence acquired). Use the default high pass filter (set to 100 s).
5. Hippocampal Seeds
- Generate a mask of the left hippocampus in each participant's high resolution structural space using FSL's FMRIB Integrated Registration and Segmentation Tool (FIRST) segmentation algorithm28.
NOTE: Other regions, including right hippocampus, would be interesting and valid seeds for further analyses. - Using a statistical software platform, write code in to calculate the length of the anterior and posterior thirds of the structure29. Specifically, use the length of the volumetric hippocampal mask in the anterior-posterior plane to find the coordinates demarking the anterior and posterior thirds of this plane.
NOTE: A recently published method of segmenting the hippocampus along the longitudinal axis might be an alternative seed creation approach30. - Based on these coordinates, create anterior and posterior hippocampal mask images. Register the anterior and posterior hippocampal masks into native functional space using the "example_func2highres" matrix in the registration directory of the FEAT output.
NOTE: Using the anterior and posterior thirds prevented signal blurring across the two hippocampal seeds after registration to functional space. There is evidence of functional specialization along the longitudinal axis of the hippocampus31,32,33,34. Anterior regions are input regions and associated with encoding, while the posterior hippocampus is an output region associated with memory retrieval and consolidation35,36,37. Thus, using these regions allows assessment of functional involvement of anterior versus posterior hippocampus in encoding versus retrieval phases of the memory task. - Use FSL mean timeseries (fslmeants) to extract the denoised average timeseries from the anterior and posterior hippocampal seeds (Figure 1). Follow the program instructions and use either the anterior or posterior hippocampal seed as the mask and the denoised, preprocessed functional data as the main image.
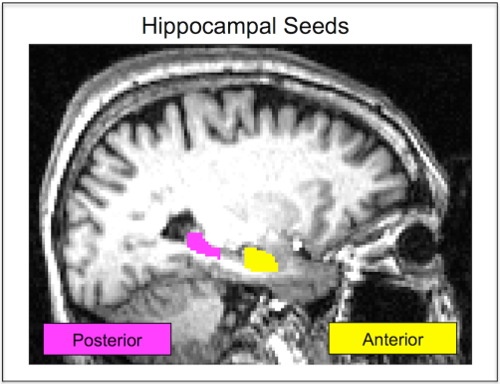
Figure 1: Hippocampal Seeds. In native space, a single participant's anterior hippocampus seed is shown in yellow. The posterior hippocampus seed for the same participant is shown in pink. Seeds are defined in each participant's unique structural image and then registered to their functional scan. Seeds are never in a standardized space, which improves the accuracy of the hippocampal segmentation. This figure has been reprinted with permission12. Please click here to view a larger version of this figure.
6. PPI Model
- Use the GUI for FSL FEAT to load the preprocessed functional data.
- In the data tab, choose the "filtered_func_data" denoised image (output from the steps completed in section 4) as the input file. In the pre-stats tab, set motion correction and brain extraction to "none." Unclick boxes to perform temporal filtering and spatial smoothing.
- PPI Model Set-Up (Table 1).
- In the stats tab, select "full model set-up". In the EVs tab, add all the EVs from the first level model: 6 motion correction EVs, confound EV matrix from FSL Motion Outliers and task timing EVs. Click the up arrow to add EVs. Include in this model an EV for the physiological timecourse from the seed (the text file output of fslmeants in step 5.4) as a covariate of no interest by clicking on the up arrow.
- Create the PPI terms.
- Choose "interaction" in the basic shape menu and select the seed timecourse EV and one task EV. For the "make zero" option, choose "mean" for the seed timecourse EV and "centre" for the task EV. Repeat this procedure for the other task phase(s). Run a separate model for each seed region.
NOTE: These new EVs are the PPI terms for the phase of the task selected (psycho) and the seed (physio). In the present study, a PPI term for the encoding phase and a second PPI term for the retrieval phase were included in each PPI model. The "centre" option ensures that the "on" and "off" phases of the block design task are treated equally. The "mean" option is always applied to the seed timecourse and results in the mean being subtracted from this regressor.
- Choose "interaction" in the basic shape menu and select the seed timecourse EV and one task EV. For the "make zero" option, choose "mean" for the seed timecourse EV and "centre" for the task EV. Repeat this procedure for the other task phase(s). Run a separate model for each seed region.
- In the contrasts and F-tests tab, model the following specific effects by entering "1" in the corresponding EV cell: psych_enc (encoding task phase), psych_ret (retrieval task phase), phys (seed timecourse), PPI_enc (PPI of seed and encoding), PPI_ret (PPI of seed and retrieval). Lastly, enter a "-1" to model negative PPIs for each task phase.

Table 1: gPPI model set-up.
7. Group Comparisons
- Select "higher-level analysis" in FSL FEAT to run a simple group model comparing APOEε4 carriers to non-carriers for each task-seed combination.
NOTE: These comparisons are run to generate the relevant group 4D residuals images ("res4d") needed to estimate the smoothness of the dataset. Statistically significant results from this group comparison are valid, but in the steps below another thresholding approach using AFNI and SPM8 to set a significant cluster minimum based on Monte Carlo simulations is described. - Use Analysis of Functional Neuroimaging (AFNI)
- Use AFNI's 3dFWHMx (any version after December 2015) at the command line to estimate the smoothness of the group 4D residuals images generated using FSL.
NOTE: A bug was discovered in AFNI's 3dClustSim and corrected in May 2015. In December 2015, AFNI's 3dFWHMx was updated to more accurately model auto-correlations. Thus, versions of these tools released in December 2015 or later should be used. - Use AFNI's 3dClustSim (any version after December 2015) to determine cluster extent minimums reaching significance at different voxel-level thresholds. Include the smoothness estimates from the previous step in the command line invocation of 3dClustSim. From the table generated by 3dClustSim, based on the study hypotheses regarding the expected effects' height and extent, choose a voxel-level threshold and corresponding cluster minimum size.
NOTE: In general, larger clusters minimize false positives.
- Use AFNI's 3dFWHMx (any version after December 2015) at the command line to estimate the smoothness of the group 4D residuals images generated using FSL.
- Use Statistical Parametric Mapping (SPM8)
- Using the SPM8 GUI, select "specify 2nd-level". The batch editor will open. Select "two sample t-test" under design. Navigate to the directory with the parameter estimate images for group 1 (APOEε4 carriers) and select by clicking on them. Next, add group 2 (APOEε4 non-carriers) images. Run this comparison by clicking on the green play button.
- Return to the SPM GUI, select "estimate", and navigate to the SPM.mat file created in the previous step to run the model estimation process.
- Select "results" and run group comparison contrasts: APOEε4 carriers > APOEε4 non-carriers, APOEε4 non-carriers > APOEε4 carriers.
- Click on "define a new contrast", choose "T-contrast" under "type" and enter "1 -1" in the "contrast" box for APOEε4 carriers > APOEε4 non-carriers. Click "done". Choose "none" for Apply Masking, and manually set the voxel-level threshold and the cluster size minimum according to the determination made in step 7.2.2.Enter "-1 1" for APOEε4 non-carriers > APOEε4 carriers.
Note: In the present study, a voxelwise threshold of p <0.005 was used and clusters thresholded at alpha <0.05.
- Click on "define a new contrast", choose "T-contrast" under "type" and enter "1 -1" in the "contrast" box for APOEε4 carriers > APOEε4 non-carriers. Click "done". Choose "none" for Apply Masking, and manually set the voxel-level threshold and the cluster size minimum according to the determination made in step 7.2.2.Enter "-1 1" for APOEε4 non-carriers > APOEε4 carriers.
With two different active task phases (encoding and retrieval) and two seed regions (anterior and posterior hippocampus) there are four conditions to report results for each group. The within-group task activation maps (not shown here, see Harrison et al., 201612) show that the occipital lobe, auditory cortex, large regions of parietal lobe, frontal language areas, superior temporal gyrus, and caudate (more pronounced during retrieval) have significant BOLD signal increases during encoding and retrieval in both experimental groups. Within-group PPI analyses revealed that there are no significant increases in functional connectivity with either anterior or posterior hippocampal seeds for either APOEε4 carriers or non-carriers. Within-group PPI analyses revealed significant decreases in functional connectivity in APOEε4 carriers for both task conditions and hippocampal subregions (Figure 2). In APOEε4 non-carriers, significant decreases in functional connectivity were only observed with posterior hippocampus during encoding (Figure 2). The positive and negative PPI maps show a divergence between APOEε4 carriers and non-carriers in how hippocampal functional connectivity changes during a memory task. To determine if the divergence is statistically significant, it is necessary to directly compare the groups for each of the four results38.
For the sake of brevity, group comparison results showing APOE-mediated differences only for one region and task phase, anterior hippocampus during retrieval, are presented here (non-carriers > carriers, Figure 3). During retrieval, the divergence of anterior hippocampus connectivity changes observed within group (Figure 2) results in significant between group differences in bilateral supramarginal gyrus, right angular gyrus and right precuneus.
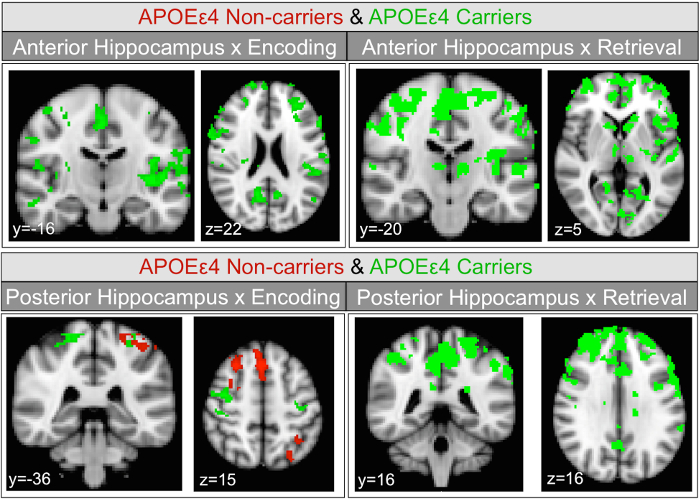
Figure 2: Hippocampal seeds task-dependent negative functional connectivity change maps. Coronal and axial views of group average task-dependent negative functional connectivity change of hippocampal subregions in APOEε4 non-carriers and carriers separately, within group. Task-dependent connectivity decreases with the anterior hippocampus seed are shown in the upper panels. The lower panels show task-dependent connectivity decreases with the posterior hippocampus. Maps were thresholded at z = 2.3, cluster corrected at p <0.05. Voxels meeting threshold in APOEε4 non-carriers (in red) and carriers (in green) are overlaid. This figure has been reprinted with permission12. Please click here to view a larger version of this figure.
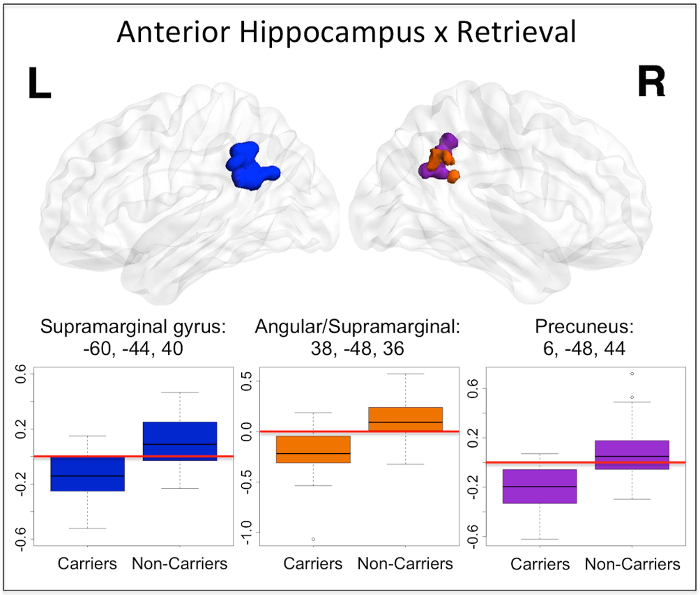
Figure 3: Anterior hippocampal seed connectivity change differences between APOEε4 carriers and non-carriers during retrieval. During retrieval, significant differences between APOEε4 carriers and non-carriers were found in left supramarginal gyrus (dark blue), right supramarginal/angular junction (orange) as well as right precuneus (purple). The results from this two-sample t-test were thresholded to reveal clusters significant at alpha <0.05 with a voxelwise threshold of p <0.005. The peak coordinate for each cluster is reported in MNI space, in x, y, z planes (mm). For illustration of the direction and magnitude of the difference between groups, contrasts of parameter estimates from each cluster are plotted by group. The red horizontal lines indicate zero and highlight that carriers have decreased (negative) functional connectivity to anterior hippocampus in these regions during retrieval. The band within the boxes represents the median while the upper and lower edges of the boxes represent the first and third quartiles, respectively. The whiskers extend up to 1.5 times the interquartile range. Data points outside this range are plotted as outliers. This figure has been reprinted with permission12. Please click here to view a larger version of this figure.
