During a standard infection experiment, mice are infected with approximately 2.5 x 108 CFU through gavage of 100 μl overnight C. rodentium culture. Infection of C57BL/6 mice with C. rodentium results in modest and transient weight loss and diarrhea. Although a rare occurrence with C57BL/6 mice, animals may become ill and require euthanization. Therefore, mice should be monitored for degree of weight loss and symptoms of distress such as piloerect fur and hunched posture, to determine the extent to which different strains are affected by the infection.
Results presented in Figures 1 through 4 are representative of an infection done using an overnight culture prepared from a frozen glycerol stock. At 7 days post-infection, mice were anesthetized and blood was collected through cardiac puncture. Figure 1 shows FITC-dextran measured in the serum of C57BL/6 mice, as well as from Toll like receptor 2 (TLR2) knockout mice, which have previously been found to exhibit impaired epithelial barrier integrity during infection9. In the presence of an intact intestinal epithelial barrier, 4 kDa FITC-dextran is poorly permeable through this layer. Therefore, increased levels of FITC-dextran in serum suggest an impairment of intestinal epithelial barrier integrity during infection allowing this molecule to leak across. As a control, serum levels in uninfected mice gavaged with FITC-dextran are also presented.
By one week post-infection, bacterial loads in the colon have been found to peak around 109 CFU/g3,4. Bacterial loads can be measured at desired time points post-infection to confirm infection, as well as to assess if different knockout mice suffer from increased or decreased bacterial burdens. As C. rodentium is a luminal pathogen that can intimately adhere to intestinal epithelial cells, bacterial loads in the luminal contents, cecal, and colonic tissues can be measured. Figure 2 shows typical bacterial loads measured at day 7 post-infection cecal and colonic tissues as well as within the intestinal lumen of C57BL/6 mice.
Most of the pathology observed during C. rodentium infection in C57BL/6 mice occurs in the distal 2 cm of the colon11. Macroscopically by day 7 post-infection, a thickening of the colonic mucosa as well as a shortening in length of the colon is observed, with little overt damage seen in C57BL/6 mice. Normally during infection, C57BL/6 mice suffer only moderate inflammation and pathology characterized histologically by immune cell infiltration, elongation of colonic crypts and goblet cell depletion (Figure 3).
As seen in the H&E sections in Figure 3, several host responses are mounted during infection with C. rodentium. To further characterize these changes, immunofluorescence staining can be utilized to examine changes in proteins of interest, such as markers of proliferation, cell death, or innate and adaptive immune responses. Immunofluorescence staining is also valuable in examining aspects of the bacterial response, such as its localization within the infected tissue. An example of this staining technique, examining a host protein ki67 (red), a marker for cell proliferation and the C. rodentium protein tir, green, to examine bacterial localization at day 12 post-infection is presented in Figure 4.
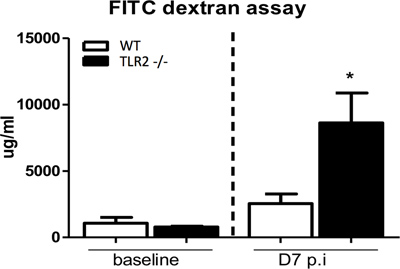
Figure 1. Measurement of serum FITC-dextran to assess epithelial barrier integrity. Mice were gavaged with 4 kDa FITC-dextran under uninfected conditions and at 7 days post-infection, following which serum FITC-dextran levels were determined. C57BL/6 (WT) mice exhibit a negligible increase in FITC-dextran serum levels at 7 days post-infection compared to uninfected conditions. In contrast, TLR2 -/- mice have significantly increased levels of FTIC-dextran in serum compared to baseline suggesting severely impaired barrier integrity in this strain during C. rodentium infection, as has previously been shown.
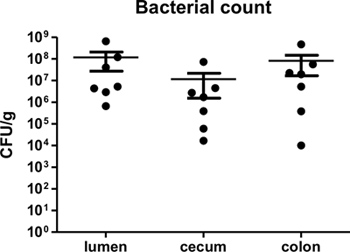
Figure 2. C. rodentium load in lumen, cecum, and colon of infected C57BL/6 mice at 7 days post-infection. Lumen, cecal, and colonic tissues were collected, homogenized, and plated in serial-dilution on LB agar. Each circle represents a single sample collected from individual animals. Solid lines indicate the geometric mean while vertical error bars indicate SEM.
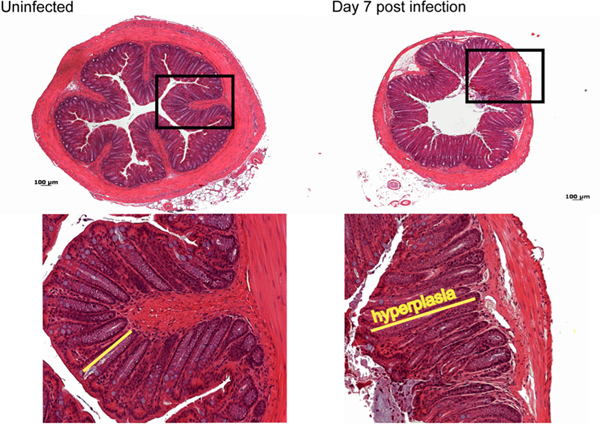
Figure 3. Histological analysis of damage caused by C. rodentium infection. By day 7 post-infection moderate immune cell infiltration, as well as elongation of crypts, goblet cell depletion and mild edema is observed compared to control uninfected tissue. Click here to view larger figure.
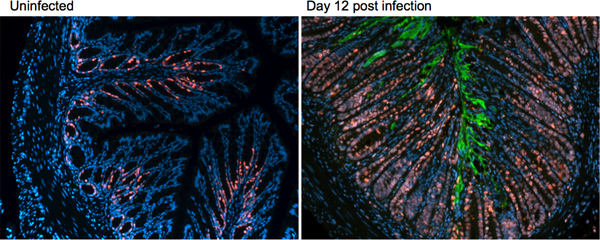
Figure 4. Immunofluorescence staining on uninfected and day 12 post-infection tissues of C57BL/6 mice. Distal colonic tissue is stained for the host protein ki67 (red) and the C. rodentium protein tir (green).
