High temperature RNA denaturation coupled to affinity purification of biotinylated cDNA prevents amplification of non-specific ASP products during PCR in PBMCs infected in vitro and in CD4+ T cells isolated from patients. RT self-priming has been shown to occur during reverse transcription of antisense RNAs10,14,15,16,17. In order to prevent this phenomenon, we developed a novel approach using the technique originally described by Heist et al.10. Our procedure involves high temperature denaturation of the RNA prior to RT and the use of biotinylated primers to perform RT, followed by affinity-purification of the biotinylated cDNAs onto streptavidin-coated magnetic beads13. The cDNAs obtained by this method can be used for downstream applications including standard PCR or qPCR.
The experimental conditions for antisense transcription were first tested in PBMCs from one healthy donor infected in vitro with HIV-1HXB2, the reference sequence for HIV-1 Subtype B (all our patients are infected with isolates of this subtype). Table 2 shows the sequences of primers and probes used in this study. Our initial RT reactions were done using a regular, non-biotinylated antisense primer (ASP R1) and resulted in the successful amplification of ASP (ASP F1-R1 primer pair) (Figure 1A, Lanes 1 – 3). However, by this approach, we also amplified a band of the same molecular weight in primer-minus RT reactions controls (Lanes 4 – 6). The complete lack of signal in our negative controls (Lanes 7 – 8) indicated that the non-specific template was coming from the RT reaction and not from cross-contamination of samples.
To bypass the problem created by RT self-priming, we decided to use a method previously described by Haist et al.10, in which the specific antisense primer is labelled with biotin so that the resulting biotinylated cDNA corresponding to the antisense orientation can be purified prior to PCR and selectively amplified. By this approach, we were able to amplify the ASP sequence (Figure 1B, Lanes 1 – 3) with greatly reduced contamination from the non-specific cDNA in the primer-minus controls (Figure 1B, Lanes 4-6).
Optimization of this method was achieved by complete denaturation of the RNA prior to RT at 94 °C, followed by immediate cooling onto iced water. As shown in Figure 2A,B, the ASP band is effectively amplified, whereas the non-specific products have disappeared.
ASP RNA is detected in CD4+ T cells from patients with detectable viraemia and in absence of therapy, following stimulation with anti-CD3/CD28. ASP RNA was undetectable in either unfractionated PBMCs or unstimulated CD4+ T cells isolated from HIV patients (data not shown). However, it could be easily detected in CD4 cells isolated from three HIV-positive subjects, MP135, MP140, and MP148 (Table 1), following stimulation with anti-CD3/CD28. The kinetics of ASP RNA expression measured by qPCR in these three patients are shown in Figure 3.
CD4 cells isolated from patients undergoing ART and with non-detectable viraemia, may produce small amounts of ASP RNA when stimulated with anti-CD3/CD28. Quantification of ASP RNA levels in two of these patients (MP071, MP146) by qPCR shows that in these conditions ASP RNA is detected in low levels (10-15 copies/million CD4+ T cells) at 3-5 days post-stimulation (Figure 4). In one patient however (MP069), no ASP RNA could be detected at any time point (data not shown).
ASP and env RNAs have similar patterns of expression in one untreated patient with detectable viraemia (MP140). Two patients were analyzed, one untreated (MP140) and one treated (MP146). In MP140 we could detect both ASP and env. Although their levels of transcription were dissimilar (Figure 5A), the expression curve of these two genes over time was identical, which can be visualized plotting data on a logarithmic scale (Figure 5B). In patient MP146, which was treated and whose viraemia was below the levels of detection, ASP and env were barely detectable and only after several days of stimulation (Figure 5C).
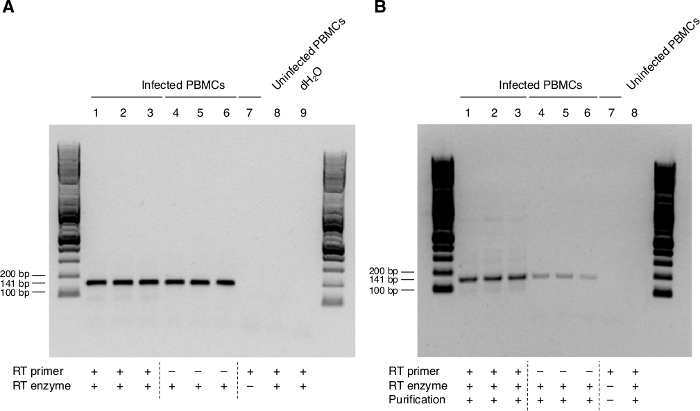
Figure 1: Expression of ASP RNA in PBMCs from one HIV-negative individual infected in vitro with HIV-1HXB2 by standard and modified RT-PCR. (A) Detection of ASP RNA using standard RT-PCR. ASP is amplified from cDNA synthesized in the presence of the non-biotinylated ASP-specific antisense primer ASP R1 (Lanes 1–3). The same band is also found in primer-minus RT controls (Lanes 4–6). (B) Visualization of ASP RNA by biotinylation of the antisense primer and cDNA purification (Lanes 1-3). A band of decreased intensity is still detected in primer-minus controls (Lanes 4–6). No bands are present in negative controls. Previously published in the article "Detection of antisense protein (ASP) RNA transcripts in individuals infected with human immunodeficiency virus type 1 (HIV-1)", by Mancarella et al.13 (J Gen Virol 100(5):863-876. doi: 10.1099/jgv.0.001244). Please click here to view a larger version of this figure.
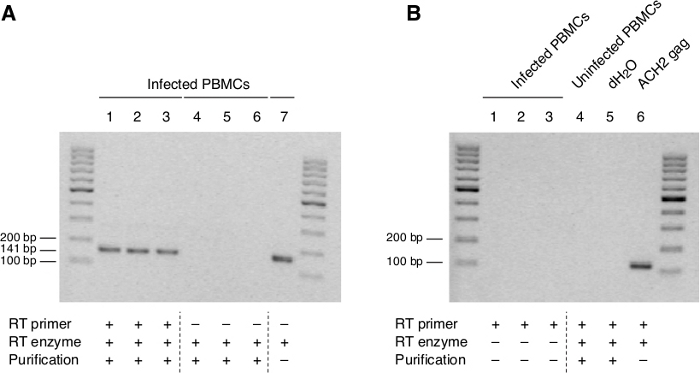
Figure 2: ASP RNA is expressed in PMBCs of one HIV-negative individual following infection with HIV-1HXB2. (A) The ASP band is easily detected RNA which has been fully denatured and reverse-transcribed using the biotinylated RT primer (ASP R1) (Lanes 1–3) but not in purified cDNA from primer-minus controls (Lanes 4–6). (B) No signal corresponding to ASP is detected in RT-minus controls of RNA from infected PBMCs, uninfected PBMCs or water, although a clear band is visible in gag positive control from ACH-2 cells. Previously published in the article "Detection of antisense protein (ASP) RNA transcripts in individuals infected with human immunodeficiency virus type 1 (HIV-1)", by Mancarella et al.13 (J Gen Virol 100(5):863-876. doi: 10.1099/jgv.0.001244). Please click here to view a larger version of this figure.
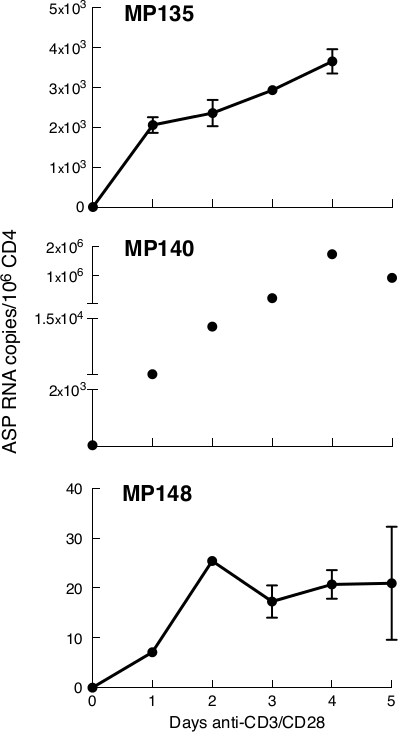
Figure 3: In anti-CD3/CD28-stimulated CD4+ T cells isolated from three untreated patients, ASP RNA production is readily detectable and peaks between day 2 and 4 post-stimulation. In MP135 and MP140, expression of ASP peaked at day 4 post-stimulation, while in MP148 it peaked at day 2. ASP levels are expressed as RNA copies/million CD4+ T cells. Points in the time course correspond to the mean value of triplicate PCR reactions. Previously published in the article "Detection of antisense protein (ASP) RNA transcripts in individuals infected with human immunodeficiency virus type 1 (HIV-1)", by Mancarella et al.13 (J Gen Virol 100(5):863-876. doi: 10.1099/jgv.0.001244). Please click here to view a larger version of this figure.
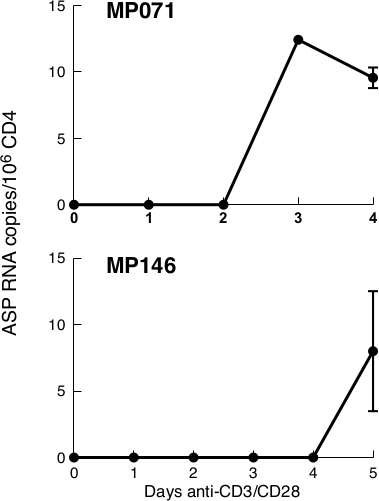
Figure 4: In two aviremic patients undergoing ART, ASP is barely detectable only after a few days of stimulation. In both our patients, ASP could not be detected at day 0. In Patient MP071, low levels of ASP could be detected at day 3, whereas in Patient MP146, we had to wait until day 5 in order to see some levels of RNA. ASP levels are expressed as RNA copies/million CD4+ T cells. Points in the time course correspond to the mean value of triplicate PCR reactions. Previously published in the article "Detection of antisense protein (ASP) RNA transcripts in individuals infected with human immunodeficiency virus type 1 (HIV-1)", by Mancarella et al.13 (J Gen Virol 100(5):863-876. doi: 10.1099/jgv.0.001244). Please click here to view a larger version of this figure.
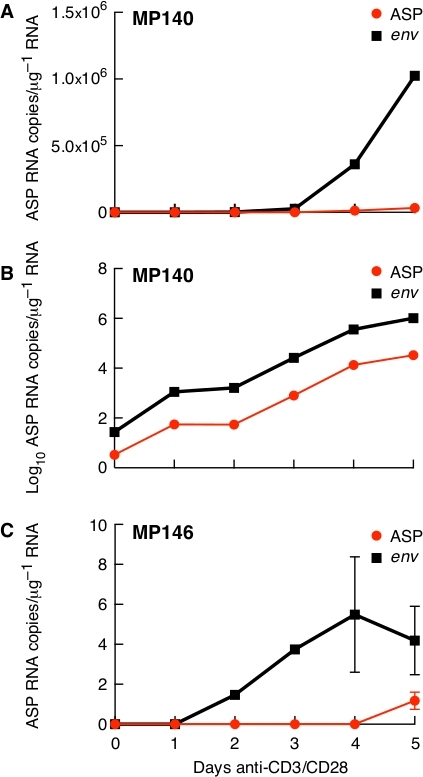
Figure 5: Expression of ASP and env in anti-CD3/CD28-stimulated CD4+ T cells in treated (MP146) and untreated (MP140) patients. (A) In the untreated patient (MP140), env is expressed at higher levels than ASP; (B) The same data plotted on a logarithmic scale show that ASP and env expression are characterized by a similar profile over-time; (C) Expression kinetics of ASP and env in one patient with undetectable viraemia and undergoing ART (MP146). Previously published in the article "Detection of antisense protein (ASP) RNA transcripts in individuals infected with human immunodeficiency virus type 1 (HIV-1)", by Mancarella et al.13 (J Gen Virol 100(5):863-876. doi: 10.1099/jgv.0.001244). Please click here to view a larger version of this figure.
| Patient ID | Age | Sex | Stage of HIV infection | Clade | Viral load (copies/ml) | CD4 Count (cells/µl) | ART status |
| MP135 | 44 | M | C3 | B | 1.6×105 | 176 | Untreated |
| MP140 | 23 | M | A2 | B | 3.6×104 | 427 | Untreated |
| MP148 | 37 | M | A1 | B | 2.0×104 | 717 | Untreated |
| MP069 | 42 | M | A1 | B | <20 | 1309 | Treated |
| MP071 | 47 | M | C3 | B | <20 | 167 | Treated |
| MP146 | 59 | M | C3 | B | <20 | 385 | Treated |
Table 1: Patients' features. Previously published in the article "Detection of antisense protein (ASP) RNA transcripts in individuals infected with human immunodeficiency virus type 1 (HIV-1)", by Mancarella et al.13 (J Gen Virol 100(5):863-876. doi: 10.1099/jgv.0.001244).
| Primer/Probe Name | Primer sequence (5' to 3') |
| ASP F1 | TTAGGAGTAGCACCCACCAA |
| ASP R1 | GAACCCAAGGAACAAAGCTC |
| PAN ASP F | ACCAAGCCTCCTACTATCATTATG |
| PAN ASP R | GCACATTGTAACATTAGTAGAGCA |
| ASP MP135, 146, 071 F | CCCAAGAACCCAAGGAACATAG |
| ASP MP135, 146, 071 R | CATTAGGAATAGCACCCACCAA |
| ASP MP135, 146, 071 Probe FAM/TAMRA | TCTCTGCACCACTCTTCTCTTTGCC |
| ASP MP140 F | CCCATAGTGCTTCCTGCTATTC |
| ASP MP140 R | AGAAGAGTGGTGCAGAGAGA |
| ASP MP140 Probe FAM/TAMRA | AGCTCCTATTGTTCCCACTGCTCT |
| ASP MP148 F | CTCTCTGCACCACTCTTCTTT |
| ASP MP148 R | AGACCTGGAGGAGGAGATATG |
| ASP MP148 Probe FAM/TAMRA | TGGTGGGTGCTACTCCTAATGGTT |
| ENV MP140 F | AGAAGAGTGGTGCAGAGAGA |
| ENV MP140 R | CCCATAGTGCTTCCTGCTATTC |
| ENV MP140 Probe FAM/TAMRA | AGCTCCTATTGTTCCCACTGCTCT |
| ENV MP146 F | CATTAGGAATAGCACCCACCAA |
| ENV MP146 R | CCCAAGAACCCAAGGAACATAG |
| ENV MP146 Probe FAM/TAMRA | TCTCTGCACCACTCTTCTCTTTGCC |
Table 2: RT-PCR primers and probes
