The study protocol received approval from the Landesdirektion Sachsen, permit numbers 25-5131/564/2, DD25-5131/450/4, 25-5131/496/56, DD25.1-5131/354/87. The zebrafish strains used were maintained according to national law and under standardized conditions as previously described27,28. The details of all the reagents and the equipment used in the study are listed in the Table of Materials.
1. Preparation of materials and solutions
- Maintain the embryos in 1x E3 media. Prepare the 1x E3 media by diluting 170 mL of a 60x E3 stock in 10 L of deionized water and adding 2 mL 0.1% methylene blue.
- Prepare the 60x E3 media stock by mixing 17.2 g of NaCl, 0.76 g of KCl, 2.90 g of CaCl2·2 H2O, 4.90 g of MgSO4·7 H2O in 1 L of deionized autoclaved water.
- For the larvae embedding, prepare aliquots of 1% low melting point agarose (LMA) in advance and keep them at room temperature.
- To prepare aliquots, dissolve LMA in 1x E3 media, e.g., 500 mg of LMA in 50 mL of 1x E3 in an Erlenmeyer flask, heat the solution in the microwave until the agarose completely dissolves, and dispense it into 1 mL aliquots in 1.5 mL tubes.
2. Zebrafish breeding and embryo collection
NOTE: Double transgenic zebrafish larvae reporting the presence of committed osteoblasts [osterix:nGFP = Tg(Ola.Sp7:NLS-GFP)zf132]29 and macrophages [mpeg:mCherry = Tg(mpeg:mCherry)gl23] were used here30 (Figure 1A).
- Set up parental zebrafish with the desired single transgenes in standard mating cages the evening before mating.
NOTE: Standard mating cages allow the eggs to pass to a lower compartment where the parental zebrafish cannot reach them.- Keep males and females separated until mating using a divider in the mating cage. Remove the divider in the morning of the next day to initiate mating.
- At noon, at the latest, remove parental zebrafish from the breeding tanks and collect the embryos, straining the water from the mating cage through a sieve.
NOTE: In case the exact age of the larvae is decisive, harvest embryos in regular intervals of approximately 30 min. Clutch mates should be used in experiments with different experimental groups. - Place the embryos in a 100 mm diameter dish containing 1x E3 media and keep them in an incubator at 28 °C31.
- Remove unfertilized eggs and dead embryos from the dish using a plastic pipette with the help of a stereomicroscope and change 1x E3 media daily.
- Before the experiment, sort larvae positive for the reporter transgenes using a fluorescence stereomicroscope.
3. Larvae embedding in low-melting agarose
- Melt LMA aliquots in a thermoblock or water bath at 65 °C. Once the LMA is melted, keep it at 42 °C until embedding (Figure 1B). Add 50 µL of 0.4% MS-222 to the melted 1 mL of LMA aliquot to achieve a 0.02% MS-222 concentration for anesthesia.
- Anaesthetize 4 dpf (days post fertilization) larvae (or older larvae) in 0.02% MS-222 E3 media.
- Transfer the anesthetized larvae to a glass-bottom microwell dish using a 3 mL plastic Pasteur pipette and remove the excess E3 media with the same pipette (Figure 1B). Avoid drying up of the larvae.
NOTE: More than one larva can be embedded in a dish simultaneously; it is recommended to embed 3 to 5 larvae to allow sufficient time for placement. - Take the melted LMA agarose from the water bath/thermoblock and add a (cooling) drop of LMA to the dish, covering the glass bottom where the larvae were positioned. Pay attention that the LMA is not too hot but still liquid.
- Then, place the larvae into the desired lateral position before the agarose solidifies. Here, ensure a lateral position in which the opercle is as close as possible to the glass bottom of the dish as a preparation for the inverse microscope setup (Figure 1B).
- Let the agarose solidify and then add E3 media (containing a maximum of 0.02% MS-222) to the dish to avoid drying of the agarose (Figure 1B).
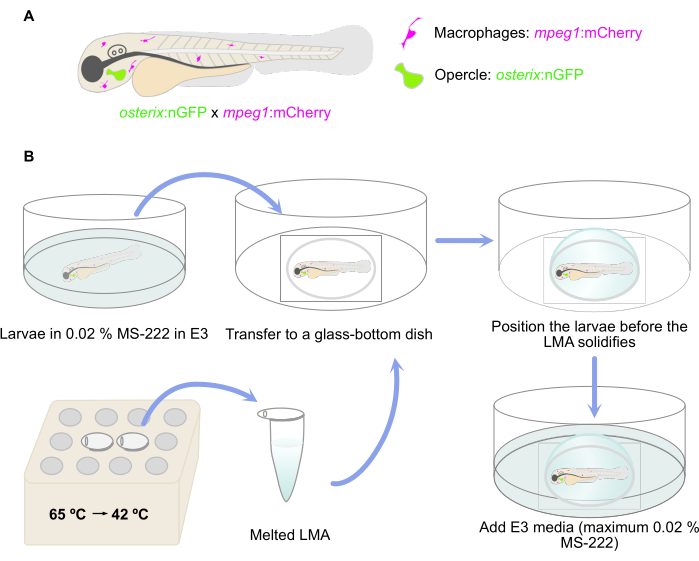
Figure 1: Schematic representation of the embedding procedure. (A) Double transgenic Tg(osterix:nGFP; mpeg1:mCherry)29,30 zebrafish larvae with green labeled opercular osteoblasts and macrophages labeled in red were used. (B) The workflow of the embedding procedure, as described in step 3 of the protocol. Please click here to view a larger version of this figure.
4. In vivo imaging of the larvae and laser ablation
- To laser-ablate osteoblasts in the developing opercle of transgenic zebrafish Tg(osterix:nGFP; mpeg1:mCherry)29,30, use an inverted two-photon laser-scanning microscope with a 25x/0.95 water objective or an equivalent setup.
- Place the dish with the embedded larvae onto the microscopic stage at the inverted microscope and position the desired region for ablation in the center of the field of view. The opercle can be easily identified with the help of the GFP fluorescence signal.
- Once the opercle is focused and centered in the field of view of the microscope, choose the z-stack settings (often 1-2 µm z-intervals are used) to image the entire breadth of the opercle. At the same time, choose the area in a specific z-plane to be ablated (usually in the posterior zone of the opercle). The two-photon laser ablation is only exerted in this plane.
NOTE: Higher resolution in the z dimension will lead to longer imaging times but will enable volumetric analyses of the acquired data later. - Image the opercle area of interest before ablation. For the GFP detection, use a white-light laser at 488 nm and a hybrid detector ranging from 495 nm to 550 nm or equivalent. Image the macrophages using 555 nm (white-light laser) excitation, with a mCherry detection range of 595 to 780 nm on a hybrid detector (or equivalent).
- Use the image format 512 x 512 for a good quality/time ratio (other formats can be used if higher resolution or alternatively higher speed are desired).
NOTE: A temperature-controlled incubator installed at the microscope can be used and adjusted, e.g., to 28 °C.
- Use the image format 512 x 512 for a good quality/time ratio (other formats can be used if higher resolution or alternatively higher speed are desired).
- Draw a circular region with a diameter of 25 µm (other shapes and sizes can be selected depending on the experiment) in a selected 2D plane, focusing the opercle using the ROI tool.
- To perform the ablation, apply ablation power (measured without objective) at 450 mW and expose the selected area to the two-photon laser at a scan speed of 400 Hz and a total duration of 10 s. During ablation, the area can be visualized using a hybrid detector in the range of 401-443 nm or equivalent.
- After ablation, confirm the ablation structurally by imaging again the same z-stack with the same settings as used prior to ablation. The ablated area does not contain fluorescent cells anymore.
5. Recovery of larvae and imaging to monitor immune cell recruitment
- If the imaging of immune cell recruitment needs to be performed with a delay, recover the larvae from the imaging and anesthesia. Remove the larvae carefully from the LMA using forceps or a dissection needle.
- First, remove a layer of agarose covering the larvae. Second, remove the agarose in direct contact with the larvae. Omit this step if immediate imaging of immune cell recruitment is taking place.
- Place the larvae back in a dish with pure E3 media and check their recovery. Recovery is evident by swimming of the larvae.
- At 4 or 6 hpl (hours post-lesion), image the opercle area again using the same settings at the same microscope to check for the effects of the ablation on macrophage recruitment. To do so, repeat the embedding (step 3) and imaging as performed before (step 4).
- After imaging, remove the larvae from the LMA. Sacrificing of the larvae can either be performed by incubation in ice water or with an overdose of MS-222 in E3, depending on the legal requirements. Specimens can then be used for follow-up studies or be disposed of.
6. Image analysis and statistical analysis
- Process the images using an image processing software such as Fiji32. Depending on the z-resolution of the acquired data, analyses can be performed in 2D or 3D.
- Quantify the macrophages manually by either including cells that are clearly separate, such as by discerning the main cell bodies, or alternatively quantifying cells with a nuclear label or by measuring the area in case cells are overlapping (next step).
- For quantification of the area, create a mask encompassing the macrophages. To do that, use the Image > Adjust > Threshold tool and create an ROI (Analyze > Tools > ROI Manager > Add) in Fiji. Exclude small particles in the background for quantification.
- Then, measure the area (Analyze > Measure). Consider whether areas of overlapping cellular fluorescence need to be included in the measurement.
NOTE: Here, the circular ablation area was omitted from the area measurements since recruited macrophages cannot be distinguished in this area.
- Then, measure the area (Analyze > Measure). Consider whether areas of overlapping cellular fluorescence need to be included in the measurement.
- Generate graphs plotting the measurement results using suitable software. To compare the data before and after ablation, perform an appropriate statistical test (here, a paired one-tailed t-test was performed).
NOTE: The figures can be prepared using photo editing and graphic design software.
Laser ablation was performed as indicated in the protocol above. The GFP signal of the osteoblasts in the ablated area disappeared instantaneously after ablation. To study the response of the osteoblast ablation in terms of the immune response, the presence of macrophages in 6 dpf larvae before and at 6 hpl was imaged. Before ablation, very few macrophages were observed in the opercle area10 (Figure 2). At 6 hpl, a strong accumulation of macrophages in the ablated opercle region and an increased number of macrophages in the field of view, including the opercle and the area surrounding the opercle, was detected10 (Figure 2).
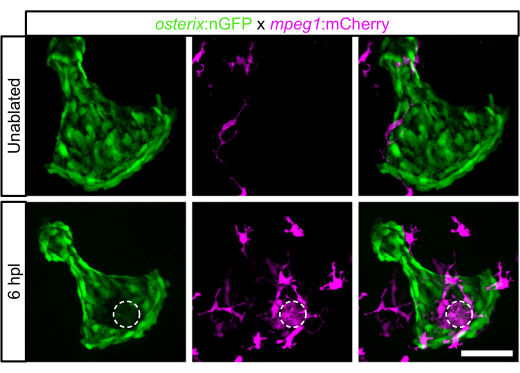
Figure 2: Macrophage recruitment at 6 hpl in 6 dpf zebrafish larvae. Opercle area without ablation and at 6 hpl. Opercular osteoblasts are labeled in green, macrophages in magenta, and the dashed circle indicates the laser-ablated area. Scale bar: 50 µm. Please click here to view a larger version of this figure.
Macrophage recruitment was also analyzed in 4 dpf larvae with much smaller opercles at 4 hpl observing similar results (Figure 3). Quantification of the macrophage number (outside the immediate laser ablation site) showed an increase within the investigated time frame (4.00 ± 2.37 macrophages before ablation versus 7.00 ± 2.76 macrophages at 4 hpl) (Figure 3B). Likewise, macrophage area (measured in µm2, 785.8 µm2 ± 723.1 µm2 before ablation versus 1553 µm2 ± 869.9 µm2 at 4 hpl) increased (Figure 3C). As macrophages are sometimes difficult to tell apart, measuring the macrophage area instead of the macrophage number is recommended, as was done here.
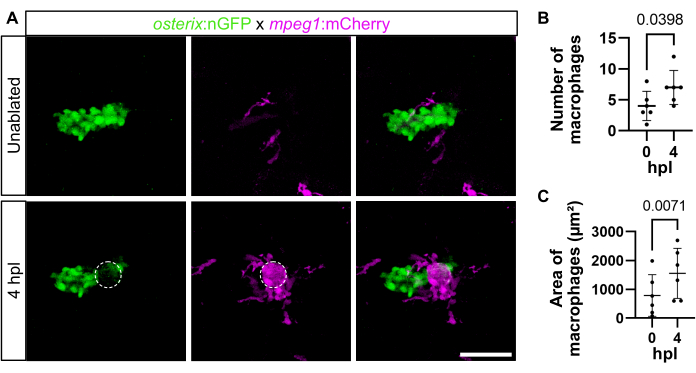
Figure 3: Macrophage recruitment at 4 hpl in 4 dpf zebrafish larvae. (A) Opercle area without ablation and at 4 hpl. Opercular osteoblasts are labeled in green, macrophages in magenta, and the dashed circle indicates the laser-ablated area. Scale bar: 50 µm. (B) Quantification of the number of macrophages in the opercle area directly before (0 hpl) and after ablation (4 hpl). (C) Quantification of macrophage area (site of laser ablation excluded). Paired one-tailed t-tests. Each dot represents the data of one larva, and the mean ± SD is shown, n = 6. Please click here to view a larger version of this figure.
