The method based on the combination of pronase E digestion and permethylation was applied to N-glycan analysis from total protein extracts of C. jejuni 11168H. Figure 1 shows a flowchart of the experimental procedure. In typical digestion, the ratio of enzyme to glycoprotein was set between 1:100 to 1:20. Here, the ratio of Pronase E to protein was increased to 2:1-3:1 and longer digestion was employed to obtain exhaust digestion by incubation at 37 °C for 48 h. The Asn-glycans from exhaust digestion were then purified by PGC and permethylated with a routine DMSO-NaOH approach. The MALDI-MS analysis of the permethylated Pronase E digests is presented in Figure 2. Three permethylation reaction times, i.e., 5 min, 10 min, and 20 min, were investigated. The obtained MS spectra indicated that the combination of Protonase E digestion and permethylation is reproducible. While the ion at m/z 1755 corresponds to Glc1GalNAc5Bac, the ion at m/z 1866 was only 111 Da higher. To verify if this ion corresponds to the permethylated product of Glc1-GalNAc5Bac-Asn, the NMR analysis of the permethylated Glc-Asn standard was performed10. The results indicated that a double bond was formed to replace the NH2 group in Asn for Glc-Asn and Glc1GalNAc5Bac-Asn. Interestingly, the data reveals the presence of free N-glycan, i.e., m/z 1755, and its relative quantity is much higher than that of Asn-linked glycan, i.e., m/z 1866. Therefore, the combination of Pronase E digestion and permethylation is not only a universal glycomics technique, but it can also distinguish free glycans from amino acid-linked glycans.
The MS and MS/MS spectra of permethylated Glc1GalNAc5Bac and Glc1-GalNAc5Bac-Asn from ESI-MS and ESI-MS/MS analysis were shown in Figure 3. The ions at m/z 1755 and m/z 1866 correspond to Glc1GalNAc5Bac and Glc1GalNAc5Bac-Asn, respectively (Figure 3A). The Glc1GalNAc5Bac-Asn structure was confirmed by annotating the MS/MS experiment (Figure 3B). Free heptasaccharide structure was further confirmed from the fragment spectrum of m/z 1755 (Figure 3C). Moreover, one can easily distinguish free N-glycans from Asn-linked glycans by observing the mass interval of 111 Da.
Figure 1: Flowchart of the experimental procedure. Please click here to view a larger version of this figure.
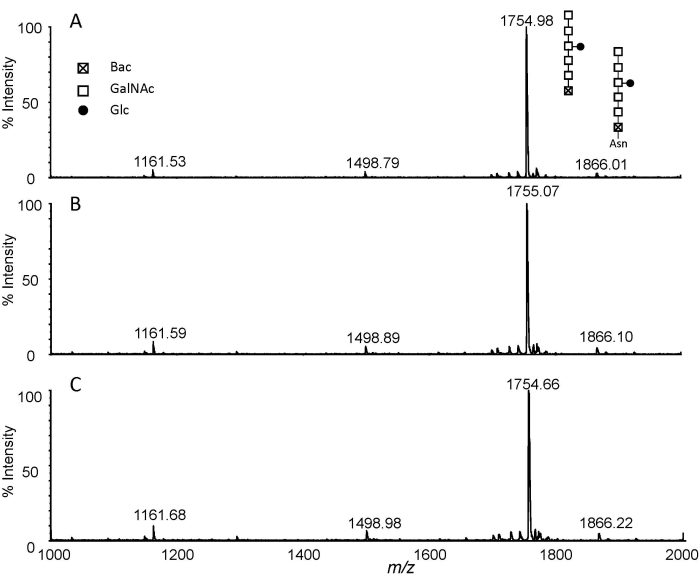
Figure 2: MALDI-TOF analysis. Extracted spectra of permethylated glycans isolated from pronase E digested C. jejuni 11168H (1 mg) with different permethylation reaction times. (A) 5 min; (B) 10 min; and (C) 20 min. Abbreviations: Glc, glucose; GalNAc, N-acetylgalactosamine; Bac, bacillosamine; Asn, asparagine. Please click here to view a larger version of this figure.
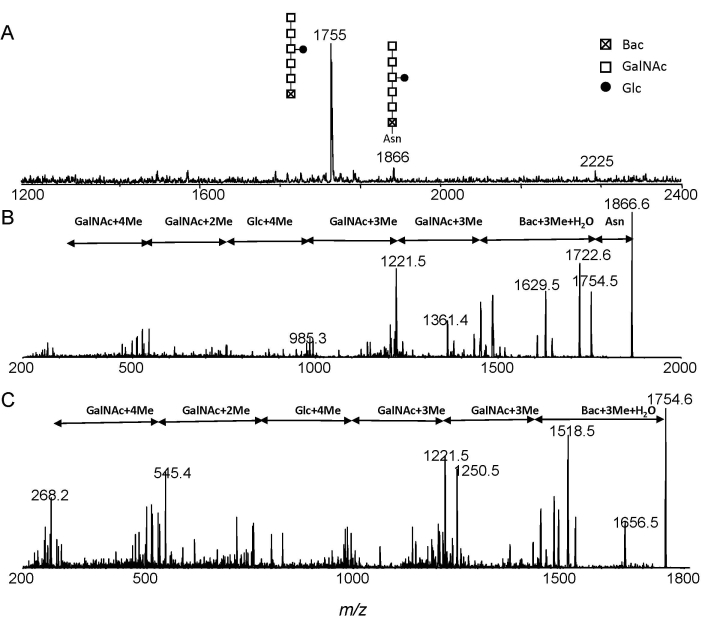
Figure 3: ESI-MS analysis. Extracted spectra of N-linked glycans from C. jejuni after Pronase E digestion. (A) Extracted MS; (B) MS/MS of precursor ion at m/z 1866; (C) MS/MS of precursor ion at m/z 1755. Abbreviations: Glc, glucose; GalNAc, N-acetylgalactosamine; Bac, bacillosamine; Asn, asparagine. Please click here to view a larger version of this figure.
