Overview of the dissection process and isolation of oral keratinocytes from the adult mouse palate
Dissociated oral keratinocytes were collected from the adult mouse palate and cultured in a customized 20% chelexed-FBS. The mouse palate consists of the hard palate and the soft palate (Figure 1C). The procedure for the isolation of mouse oral keratinocytes is summarized in Figure 1D. The palate tissue is dissected and transferred to a media containing an antibiotic-antimycotic solution before being incubated in 0.025% trypsin solution at 4 °C overnight. The following day, the palate tissue is treated with trypsin inhibitor solution and complete culture medium in equal volumes. Subsequently, the tissues are scraped using a surgical scalpel blade to collect oral keratinocytes. The cell suspension is filtered through a 100 µm cell strainer and centrifuged. The cells are then seeded in Collagen I-coated 24-well plates containing 2 mL of complete culture medium + chelexed-FBS.
Representative results of the successful isolation of mouse oral keratinocytes
Primary oral keratinocytes grew as a monolayer and displayed a cobblestone morphology (Figure 2). Small keratinocyte colonies were visible at 3-5 days (Figure 2A,B); these grew larger and formed tight colonies at 1 week of incubation (Figure 2C). Keratinocyte colonies displayed the typical morphological features of basal keratinocytes, indicating their healthy conditions. Human oral keratinocytes remained undifferentiated for several passages in the complete culture medium containing 0.06 mM Ca2+16,28. The first passage was performed approximately 2 weeks from the initial plating (Figure 2D). At later passages, keratinocytes exhibited stable growth with a shorter period of culture (Figure 2E–G). Keratinocytes stopped growing if significant fibroblasts contamination occurred during the isolation process (Figure 2H).
Isolated mouse oral keratinocytes express basal cell markers
To confirm the status of primary oral keratinocytes, immunostaining was performed using the basal cell markers Keratin 14 (K14) and α6-integrin34. K14 and α6-integrin were expressed in keratinocytes after culturing (Figure 3A,B). The cells were also stained with stem cell marker p63 to confirm their stemness. Early passage (passage 4) and late passage (passage 7) cells showed uniform expression of p63 (Figure 3C,D). In contrast, keratinocytes treated with high calcium (1.2 mM induction for 2 days) exhibited decreased p63 expression (Figure 3E), indicating that high calcium treatment suppresses stem cell-related genes in primary keratinocytes as previously reported16. The differentiation marker Keratin 13 (K13) showed rare or no expression in both early and late passages and significant expression under high calcium treatment (Figure 3F–H). To test the possibility of fibroblast contamination in the keratinocyte culture, staining using the fibroblast marker PDGFRα was performed with the same set of keratinocytes compared with mouse embryonic fibroblast (MEFs). There was no expression of PDGFRα in the keratinocyte culture, compared with the high expression observed in MEF cells (Figure 3I–3L). These results indicated that this protocol could successfully isolate basal keratinocytes and maintain these cells in the undifferentiated state.
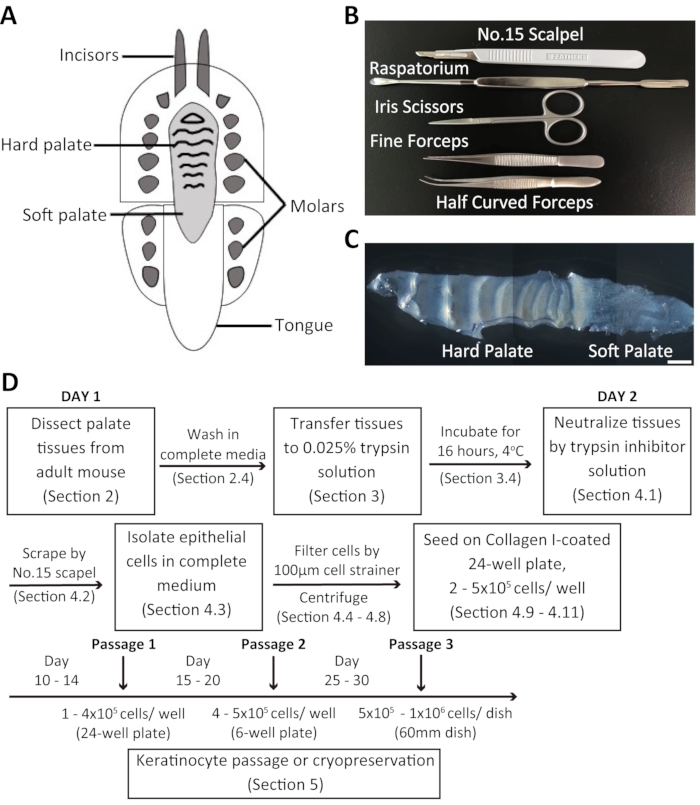
Figure 1: Overview of the dissection procedure and isolation of mouse oral keratinocytes. (A) Schematic representation of the mouse oral cavity. (B) Instruments used to dissect the palate and isolate mouse oral keratinocytes. (C) Brightfield image of the mouse palate. Scale bar: 100 µm. (D) Summary of the protocol. Please click here to view a larger version of this figure.
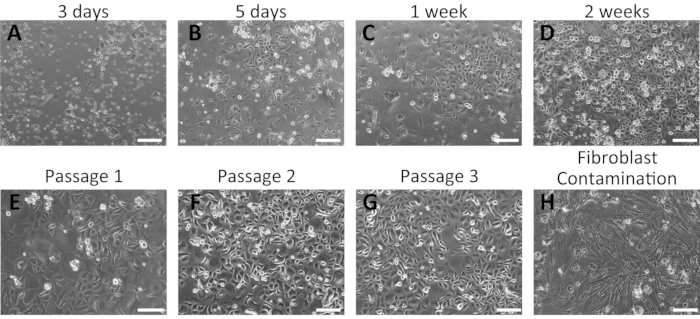
Figure 2: Representative results of the successful isolation of mouse oral keratinocytes. (A–G) Time-course images of cultured primary oral keratinocytes at 3 days (A), 5 days (B), 1 week (C), and 2 weeks (D) of culture after isolation. Morphologies of mouse oral keratinocytes after the first (E), second (F), and third (G) passages are shown. (H) Example of fibroblast contamination in mouse oral keratinocyte culture. Scale bar: 400 µm. Please click here to view a larger version of this figure.
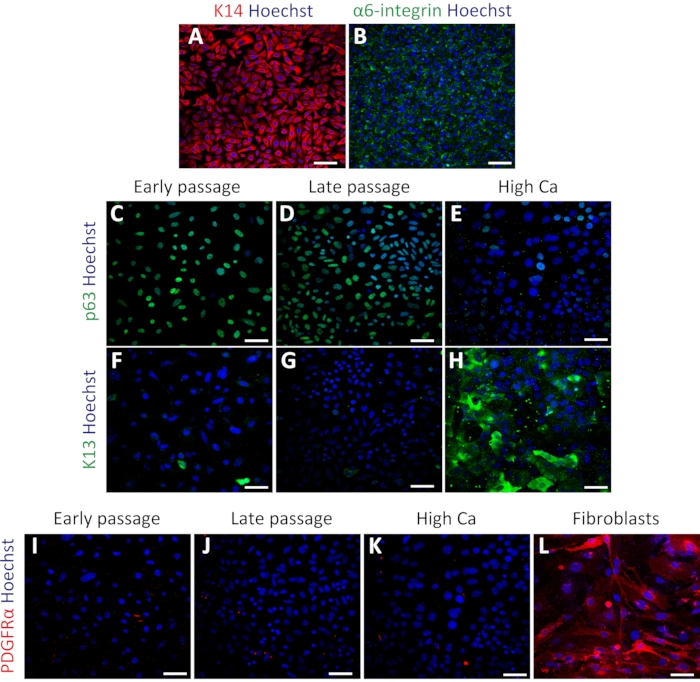
Figure 3: Isolated mouse oral keratinocytes express basal cell markers. (A–B) Representative images of immunofluorescent staining of K14 (A; red) and α6-integrin (B; green) in passage 4. (C–E) Representative images of immunofluorescent staining of p63 (green) in passage 4 (C), passage 7 (D), and high calcium treatment (E). (F–H) Immunostaining images of K13 (green) in passage 4 (F), passage 7 (G), and high calcium treatment (H). (I–L) Immunostaining images of PDGFRα (red) in passage 4 (I), passage 7 (J), high calcium treatment (K), and MEFs (L). Nuclei are stained with Hoechst (blue). Scale bars: 100 µm. Please click here to view a larger version of this figure.
