Surface preparations of the cochlear epithelium, in combination with immunolabeling and confocal imaging, have been used broadly in hearing science for the investigation of cochlear pathologies, such as quantification of ribbon synapses, sensory hair cells, and protein expression in sensory hair cells5,6,7,8. Although the dissection of adult mouse cochleae for surface preparations is not simple, new graduate students are able to learn this modified method after practicing with 10–15 ears (Figure 1 and Figure 2). With this technique, in combination with immunolabeling for CtBP2 (a marker for presynaptic ribbons) and GluA2 (a marker for post synaptic terminals), counting IHC/auditory nerve synapses using confocal images under Z projections with 0.25 µm intervals, based on the size of mouse ribbon synapses, is possible21. This is consistent with published results4,6,13. Surface preparations of CBA/J mice (without treatment) at the age of 10–12 weeks immunolabeled with CtBP2 (red) and GluA2 (green) showed that both presynaptic ribbons and post synaptic terminals are located below the IHC nuclei and are juxtaposed, indicating functional synapses (Figure 3)6,13.
By immunolabeling surface preparations with different antibodies, assessment of molecular signaling and structure in sensory hair cells is possible. For example, Figure 4 shows the results for immunolabeling for myosin VIIa and counterstaining with phalloidin and 4’,6-diamidino-2-phenylindole (DAPI). Parallel immunolabeling experiments with and without the cell and tissue adhesive using identical solutions have been conducted to assess if the adhesive used interferes with the immunoreactions. Cochlear surface preparations were immunolabeled with myosin VIIa and counterstained with phalloidin. Confocal images were taken with a 63x magnification lens under identical conditions and equal parameter settings for laser gains and photomultiplier tube (PMT) gains. There was no difference in immunoreactions or uniformity with and without the cell and tissue adhesive (Figure 5). Using different fixatives, surface preparations have provided the basis for scanning electron microscopy (SEM) images for visualization of cochlear stereocilia9. C57BL/6J mice (without treatment) at the age of 6–8 weeks show well-organized V-shaped stereocilia of OHCs in three rows (Figure 6). Additionally, cochlear surface preparations have been used to determine the pattern of expression of a report gene (i.e., GFP) and confirm successful transduction and identify transduced cell types.
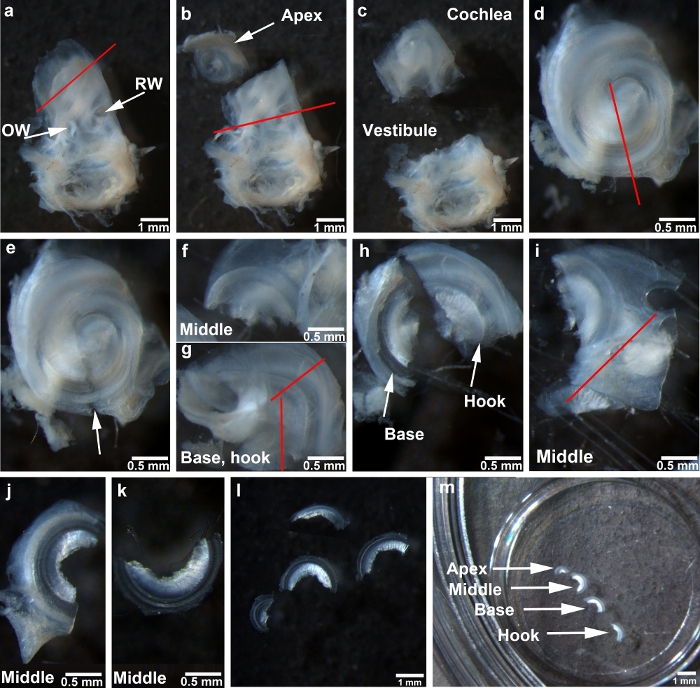
Figure 1: Depiction of the steps for adult mouse surface preparations. (a) The cochlear bony capsule of the temporal bone faces up. The red line indicates the first cut to separate the apical turn. RW = round window, OW = oval window. (b) The apex is separated from the cochlea as indicated by the arrow. The red line between round window and oval window indicates the second cut made in the cochlea. (c) The cochlear portion that contains the middle, basal, and hook regions is separated from the vestibular portion. (d) The cochlear portion is situated facing up. The red line indicates the third cut, directed towards the end from which the apical section was removed. (e) The arrow indicates the gap left after the completion of the third cut. (f) This image shows the middle region. (g) This image shows the combined basal and hook regions. The vertical red line indicates the cut between the modiolus and hook region. The cut between the basal and hook regions is indicated by the other red line. (h) Basal and hook regions are separated as indicated by the arrows. (i) The middle region of the bony capsule is cut as indicated by the red line. (j) The image shows the middle region after one side of the bony capsule is removed. (k) This image shows the middle region after completion of dissection. (l) All four regions are completely dissected. (m) Cochlear turns affixed to a 10 mm round coverslip transferred to a four-well Petri dish with the four regions as indicated. All images were taken under a stereo-dissection microscope at 1.2x, 2.5x, and 0.6x magnifications. Scale bars are indicated in each image. Please click here to view a larger version of this figure.
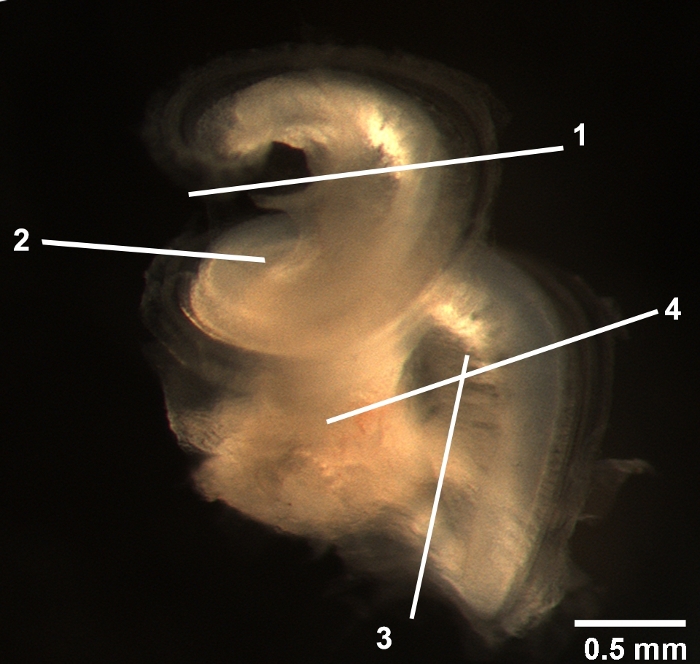
Figure 2: Location of the major cuts. The entire cochlear bony capsule was removed. The locations of the major cuts are indicated. 1) The location where the apical turn is cut. 2) The site where the middle turn is separated. 3) The critical cut to remove the modiolus. 4) The line to divide the basal and hook regions. Scale bar = 0.5 mm. Please click here to view a larger version of this figure.
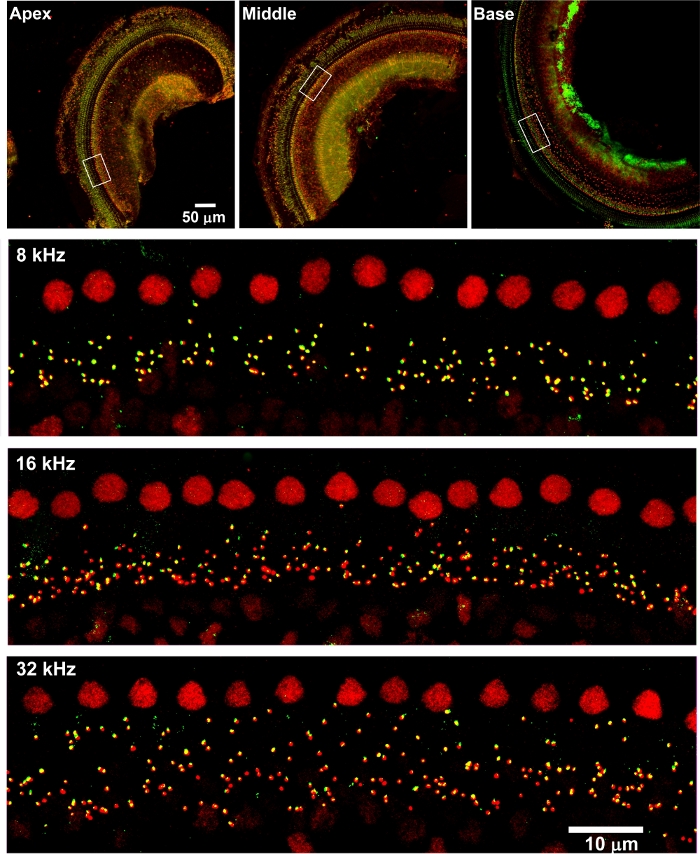
Figure 3: Confocal images reveal immunolabeling for CtBP2 and GluA2 on adult mouse surface preparations. The apical, middle, and basal regions are labelled. The lower magnification confocal images were taken with a 10x lens. Red = CtBP2, green = GluA2. Scale bar = 100 µm. Confocal images of 5-, 16-, and 32-kHz regions were taken with a 63x lens. Enlarged views of the areas indicated by the white rectangles in the apex, middle, and base portions. Scale bar = 10 µm. Please click here to view a larger version of this figure.
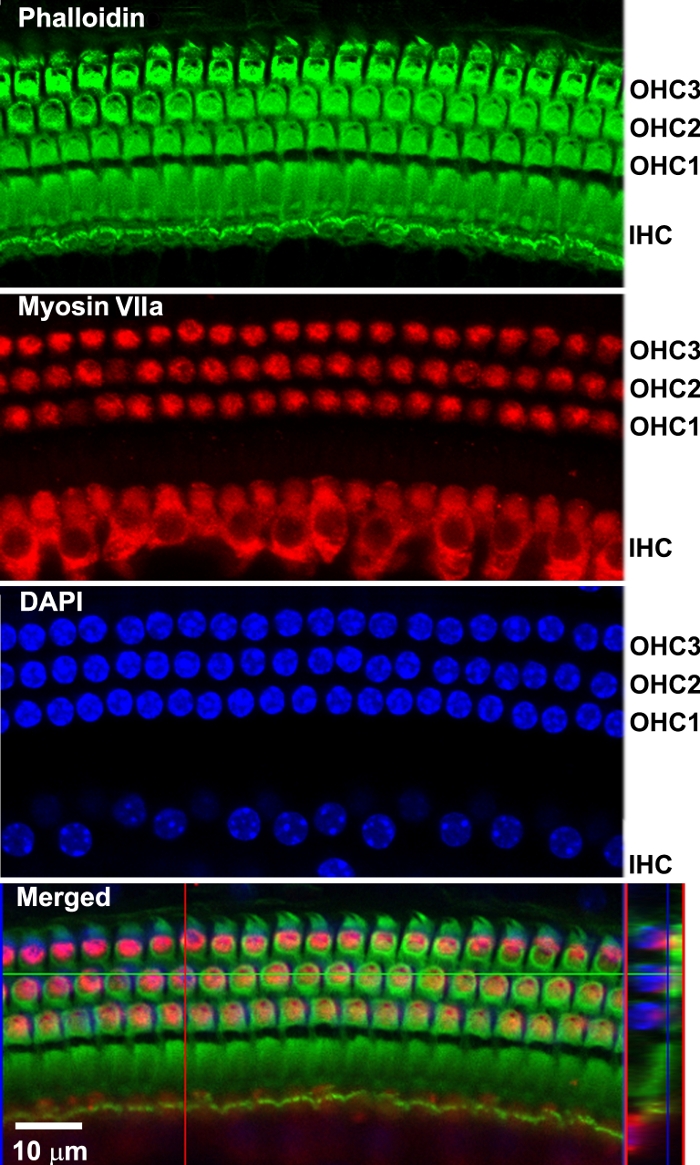
Figure 4: Confocal images reveal immunolabeling of sensory hair cells of a surface preparation from the 32-kHz region. All images were merged Z-stack projections. Phalloidin (green) stained the structure of the three rows of OHCs and one row of IHCs. Myosin VIIa (red) immunolabeled three rows of OHCs and one row of IHCs. DAPI-stained hair cell nuclei. Merged confocal images were reconstructed for side views of sensory hair cells. Scale bar = 10 µm. Please click here to view a larger version of this figure.
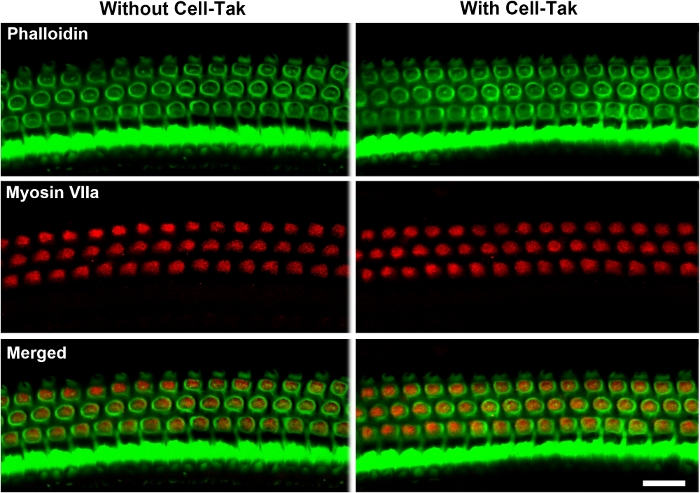
Figure 5: Cochlear surface preparations with and without the cell and tissue adhesive were processed in parallel, immunolabeled for myosin VII (red), and counterstained with phalloidin (green) using identical solutions. The cell and tissue adhesive used does not interfere with immunoreactions. Confocal images were taken with a 63x magnification lens under identical conditions and equal parameter settings. Images were taken from the 32 kHz region. Immunolabeling for myosin VIIa and staining for phalloidin in OHCs was similar with and without the cell and tissue adhesive. Scale bar = 10 µm. Please click here to view a larger version of this figure.
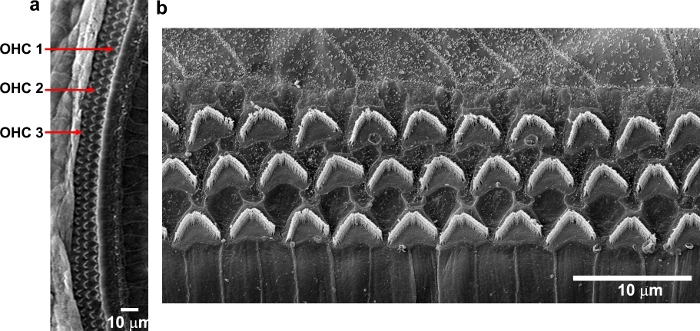
Figure 6: Scanning electron microscopy shows three rows of OHC stereocilia from C57BL/6J mice. Images were taken from the middle region. (a) A low-magnification view of three rows of OHCs. (b) Enlarged images show three rows of well-organized OHC stereocilia that appear in “V” shapes. Scale bar = 10 µm. Please click here to view a larger version of this figure.
| Name | ||||||||
| Distance from the apex (mm) | 0.4 | 1 | 2.4 | 3.3 | 3.9 | 4.7 | 5.7 | |
| Distance from the apex (%) | 7.7 | 18 | 43 | 54 | 68 | 82 | 100 | |
| Frequencies (kHz) | 6 | 8 | 16 | 22 | 32 | 48 | ||
Table 1: Mapping of the CBA/J mouse cochlear frequency sensitivity as a function of distance from the apex according to Muller1 and Viberg and Canlon2.
