Live imaging of optogenetic and non-optogenetic TDP-43 proteins in the mnr2b+ spinal motor neurons of zebrafish larvae
To induce TDP-43 phase transition in the spinal motor neurons in zebrafish, a human TDP-43h that is tagged with mRFP1 and CRY2olig22 at the N- and C-termini, respectively, was constructed and designated as opTDP-43h14 (Figure 1A). The opTDP-43h gene fragment was introduced into a BAC containing the mnr2b locus (Figure 1B). The resulting BAC, designated as mnr2b-hs:opTDP-43h, was introduced into the zebrafish genome by Tol2 transposon-mediated BAC transgenesis19. To monitor the localization of non-optogenetic TDP-43 in the spinal motor neurons, a zebrafish TDP-43 encoded by tardbp gene was tagged with EGFP at the N-terminus (Figure 1A) and the EGFP-TDP-43z gene fragment was introduced into the mnr2b BAC, similar to opTDP-43h (Figure 1B). The resulting BAC, designated as mnr2b-hs:EGFP-TDP-43z, was introduced into the zebrafish genome by Tol2 transposon-mediated BAC transgenesis. The fish injected with each BAC construct was crossed with a wild-type fish and the resulting F1 fish were screened on day 3 for red (Tg[mnr2b-hs:opTDP-43h]) or green (Tg[mnr2b-hs:EGFP-TDP-43z]) fluorescence in the ventral spinal cord. The isolated red or green fluorescence-positive F1 fish were designated as Tg[mnr2b-hs:opTDP-43h] or Tg[mnr2b-hs:EGFP-TDP-43z], respectively.
Tg[mnr2b-hs:opTDP-43h] Tg[mnr2b-hs:EGFP-TDP-43z] double-transgenic fish at 48 hpf were anesthetized and embedded in a low melting agarose on their side; after agarose solidification, they were covered with E3 buffer to allow observations of the lateral side of the spinal cord from above. Confocal laser scanning microscopy was performed with a 20x water immersion objective lens and a combination of excitation/emission wavelengths: 473/510 nm for EGFP and 559/583 nm for mRFP1. The scanned fish was immediately and carefully taken from the agarose, transferred into E3 buffer in a six-well plate, and illuminated by a blue LED light by placing the plate on a LED panel at 28 °C (Figure 2A,B). After a 24-h illumination, the fish was anesthetized and embedded again in agarose before being scanned with a confocal microscopy using the same imaging conditions, except that the ROI was adjusted to include the corresponding spinal cord region imaged at 48 hpf (Figure 2B). The whole hemispinal cord of 4-5 contiguous spinal segments was imaged during the microscopy session (typically 20-30 min), generating z-series images containing 20-40 slices depending on the horizontalness of the longitudinal axis of the spinal cord of the mounted fish relative to the confocal scanning plane.
Visualization of cytoplasmic relocation of optogenetic TDP-43 in spinal motor neurons
To visualize the cytoplasmic relocation of opTDP-43h in single spinal motor neurons, the z-series images (typically 20-40 slices) acquired at 48 and 72 hpf were opened with Fiji and separated into each EGFP-TDP-43z and opTDP-43h channel. Max intensity projection images of EGFP-TDP-43z were created for images at 48 and 72 hpf and a single isolated spinal neuron identifiable in both images at 48 and 72 hpf was selected (Figure 2B, arrows). Spinal motor neurons with cell bodies located on the dorsal side of the motor column were considered suitable for measurements of the cell body shape because of their sparse distribution patterns.
After adjusting the intensity of the EGFP signal in EGFP-TDP-43z images, ROIs covering the somas of the motor neurons were set by tracing the edge of the EGFP signal at 48 and 72 hpf (Figure 2C). The fluorescent intensity along the major axis of the soma was measured for the EGFP-TDP-43z and opTDP-43h signals at 48 and 72 hpf (Figure 2C, right). At 48 hpf, the patterns of opTDP-43h and EGFP-TDP-43z signals largely overlapped each other.
In contrast, at 72 hpf, the peak of the opTDP-43h signal was found in the region where the EGFP-TDP-43z signal was low, namely in the cytoplasm, indicating the cytoplasmic relocation of opTDP-43h. The light-dependent cytoplasmic opTDP-43h relocation is initiated largely independently of non-optogenetic TDP-4314. In this assay, Max intensity projection images were used to estimate the position of the nucleus/cytoplasm boundary at the expense of quantitative measurements.
Ratiometric comparison of the fluorescence intensity of optogenetic and non-optogenetic TDP-43 in the spinal motor neurons
To evaluate the effects of the blue light stimulation on the fluctuating opTDP-43h protein levels, opTDP-43h signals in the cell body were measured before and after the light stimulation with reference to the EGFP-TDP-43z signal. The z-series images, including the spinal hemisegment, were opened with Fiji. To set ROIs covering single spinal motor neurons, Max intensity projection images of the EGFP-TDP-43z signal were created for 48 and 72 hpf, and single isolated spinal neurons that were identifiable in both 48 and 72 hpf images were selected (Figure 3A). Using freehand selections, ROIs were drawn and registered in a ROI manager for each of the 48 and 72 hpf images (Figure 3A). Max intensity projection of the EGFP-TDP-43z signal was considered suitable to set the ROIs because of its high contrast.
To quantify the opTDP-43h and EGFP-TDP-43z signals, Sum slices projection images for each channel of the same z-series images were produced for 48 and 72 hpf (Figure 3A). Using the registered ROI sets, the fluorescent intensities of opTDP-43h and EGFP-TDP-43z were measured for each time point. Relative amounts of opTDP-43h to EGFP-TDP-43z (RFP/GFP values) were calculated for each cell and time point by dividing the RFP value by the GFP value (Figure 3B). Of the four mnr2b-positive cells that were investigated, cell #1 notably displayed a saturated EGFP-TDP-43z signal (Figure 3A). Cells with saturated fluorescent signals should be excluded from the data sets when conducting quantitative analyses. Cells #2-#4 displayed decreasing trends of relative opTDP-43h levels to EGFP-TDP-43z after blue light illumination (Figure 3B), although a larger-scale analysis previously demonstrated that the decrease in relative opTDP-43h levels to EGFP-TDP-43z was not statistically significant (65 cells from three independent animals)14.
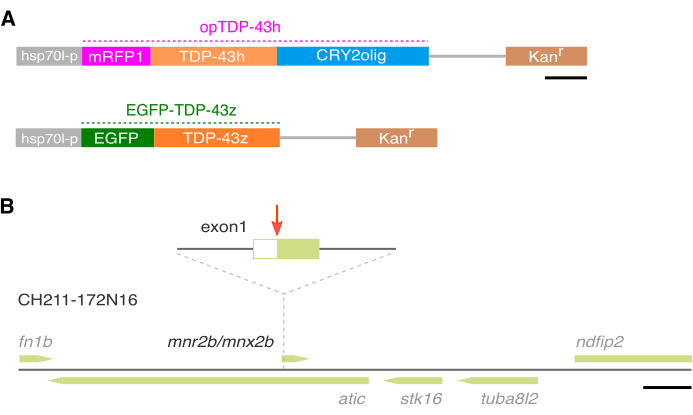
Figure 1: Construction of BAC DNA using the mnr2b locus. (A) Structures of the expression cassettes for opTDP-43h and EGFP-TDP-43z. (B) The zebrafish genomic region carried by the CH211-172N16 BAC DNA. The PCR-amplified expression cassettes for opTDP-43h and EGFP-TDP-43z are inserted downstream of the 5′-untranslated region (UTR) of the mnr2b (red arrow) in the first exon of mnr2b by homologous recombination. The bars indicate 500 (A) and 10k (B) bp. Please click here to view a larger version of this figure.
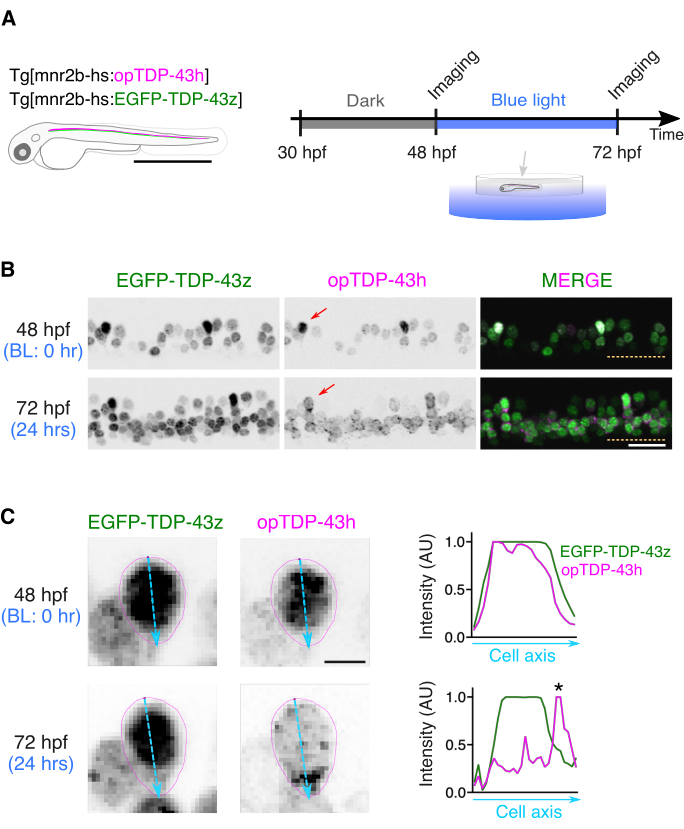
Figure 2: Live imaging of opTDP-43h before and after light stimulation. (A) A scheme of the light stimulation of opTDP-43h expressed in the spinal motor neurons of unrestrained Tg[mnr2b-hs:opTDP-43h] Tg[mnr2b-hs:EGFP-TDP-43z] double-transgenic fish through a field illumination of blue LED light from 48 to 72 hpf. (B) Max intensity projection images of the z-series confocal images of the ventral spinal cord before (48 hpf) and after (72 hpf) the light stimulation. The dashed lines demarcate the ventral limit of the spinal cord. Figures are adapted from Asakawa et al.14 (C) Cytoplasmic mislocalization of opTDP-43 after the 24-h blue light illumination. The contours of the soma of an mnr2b-positive cell (red arrows in B) observed at 48 and 72 hpf are shown in magenta. The major axes of the somas were shown with light blue. The normalized fluorescent intensity along the major axes was plotted. The asterisk indicates a cluster of strong opTDP-43 signals that do not display a strong EGFP-TDP-43z overlapping signal, indicating cytoplasmic opTDP-43h mislocalization. The bars indicate 1 mm (A), 20 µm (B), and 4 µm (C). Please click here to view a larger version of this figure.
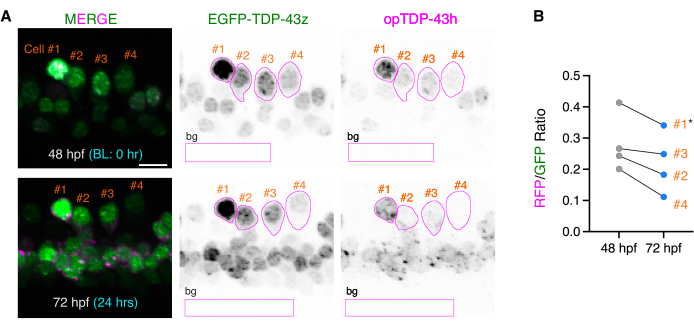
Figure 3: Ratiometric comparisons of opTDP-43h and EGFP-TDP-43z before and after light stimulation. (A) ROIs covering the somas of four single mnr2b-positive cells at 48 and 72 hpf were drawn based on the EGFP-TDP-43z signal and are shown in magenta. The rectangular ROIs (bg) were used to subtract the background signal (background ROI). Figures are adapted from Asakawa et al.14 (B) The relative intensities of opTDP-43h to EGFP-TDP-43z were plotted for each cell at each time point as the RFP/GFP ratio. Cell #1, indicated by the asterisk, was not suitable for the ratiometric comparison because its EGFP-TDP-43z signal was saturated and its RFP/GFP value was overestimated. The bar indicates 10 µm. Please click here to view a larger version of this figure.
