1. Pré-requisitos e Equipamentos de Laboratório NOTA: O conjunto da câmara de medição é descrito na Figura 3. A sanduíche de concepção da câmara de medição é constituído por três componentes principais: uma lâmina de vidro de quartzo (sílica fundida), uma película de vedação e uma lamela que veda a câmara de fluxo. A câmara de medição é montado sobre um suporte de amostra de personalização. As dimensões da câmara de amostra e suporte metálico são voltadas para atender a uma lâmina de microscópio de vidro de quartzo padrão (76 mm x 26 mm). Corte de quartzo lâminas de vidro usando uma broca de diamante (0,75 mm) nas posições indicadas na Figura 4. A concepção das lâminas de vidro de quartzo é assimétrica, a fim de diferenciar entre os dois lados de cada lâmina. NOTA: As lâminas de quartzo pode ser re-utilizado depois da medição até que a superfície se torna riscada. Para montar as câmaras, usar suportes de amostra de metal personalizado como representado na Figura 5 </ Strong>. Os suportes de amostras conter dois fios (M4), a fim de ligar uma entrada e uma tubagem de saída para a câmara de fluxo. Além disso, a utilização de fios (M3), para montar a câmara da amostra no suporte de metal, bem como roscas (M3) para fixar o suporte de prisma para a metade inferior do suporte de metal. Realizar as medições smFRET em um tipo de prisma total de microscópio reflexão interna de fluorescência (TIRFM) (Figura 6). NOTA: O TIRFM apresenta três lasers: um verde (532 nm, Nd: YAG) e um laser vermelho (643 nm, diodo laser) para a excitação do dador e as moléculas de corante aceitador, bem como um laser azul (491 nm , laser de estado sólido bombeado-diodo) para o branqueamento de fundo impurezas fluorescentes na câmara da amostra antes da medição smFRET. Os três feixes de laser são espacialmente combinado e pode ser seleccionado através de um filtro sintonizável acústico-óptico (AOTF). A luz de fluorescência é coletada por um objectivo elevado abertura, dividido em um doador e um acceptou canal, utilizando um espelho dicróico e projetadas em duas câmeras EM CCD. A câmara da amostra é ligado a uma fase micrómetro permitindo o movimento em x e y-sentido com dois motores de passo. Um terceiro motor piezo é usado em conjunto com um IR-laser e um detector sensível à posição para construir um sistema de auto-foco para assegurar foco ideal durante todo o experimento. Use alternada de excitação laser (ALEX) quando a dinâmica nas trajetórias de tempo FRET são observados 25. Essa dinâmica pode ser causada por qualquer mudanças conformacionais dentro das moléculas ou por flutuações na luminosidade receptor e piscando aceitador. NOTA: ALEX permite a discriminação entre essas duas causas possíveis e evita erros de interpretação das trajetórias FRET dinâmicos. No entanto, por razões de simplicidade, a parte do protocolo é restringido à análise dos filmes produzidos sem o Alex. Cuidado: lasers de classe 3B são usados na configuração de fluorescência única molécula. Garantir que adequada laprecauções de segurança ser, de acordo com os regulamentos governamentais locais, foram tomadas antes que o sistema é operado. Realizar a medição de absorção utilizado para a determinação do rendimento quântico de um espectrómetro de UV-VIS (ver Materiais e Métodos). Executar a medição do espectro de emissão de fluorescência de dador, o espectro de absorção de aceitador e a anisotropia de fluorescência num espectrómetro de fluorescência (ver Materiais e Métodos). Prepare câmaras de amostras de acordo com procedimentos publicados 33. Alternativamente, o procedimento descrito em [34] pode ser usado. Rotular as amostras investigadas com um dador-aceitador molécula corante par adequado para smFRET e assegurar que há uma porção de biotina para a imobilização na superfície da câmara de amostras. NOTA: Para localizar uma posição corante antena desconhecido com o software Fast-NPS são necessários diferentes construções de amostra. Cada construção needs ter uma etiqueta na posição de corante antena desconhecido e uma etiqueta numa posição de satélite conhecido a partir da estrutura de cristal. Pelo menos três construções diferentes com corantes ligados à posição da antena de satélite e três posições diferentes são requeridas para resultados precisos. Medidas entre antenas, bem como entre os satélites também são úteis para melhorar a precisão, no entanto, isso requer troca de moléculas de corante que precisa ser corretamente inseridos na análise. 2. Montagem do fluxo de Chambers em um suporte personalizado Puxe tubo de silicone (0,8 mm de diâmetro, 2,4 mm de diâmetro externo) para parafusos guia ocos (M4) e cortar o tubo em ambas as extremidades retas deixando uma saliência de 1 cm em ambos os lados, usando uma lâmina afiada. Ajustar a saliência do tubo de cerca de 2 mm no lado do parafuso de guia. Montar a câmara de fluxo no suporte da amostra de uma maneira que os orifícios da corrediça de vidro de quartzo combinar os fios no suporte da amostra. Apertarorifícios de entrada e de saída de parafusos com cuidado para garantir que a entrada e a saída da câmara de amostra são ainda penetrável. Suavemente aperte os quatro parafusos do suporte de vidro acrílico para fixar a posição da câmara de fluxo. tubagem cortada de silicone (0,58 mm ID, 0,96 mm de diâmetro externo) dentro de 20 centímetros de comprimento. Insira uma das peças para a entrada e o parafuso de saída da câmara de medição. Fechar o tubo de entrada e de saída, usando uma pinça. NOTA: As câmaras de amostras montadas pode ser armazenado à temperatura ambiente até duas semanas. 3. smFRET de medição em TIRF Microscópio Usar uma seringa, para lavar a câmara de amostras com 500 uL de PBS. Impedir as bolhas de ar que entra na câmara de amostra em todos os momentos através da criação de uma gotícula no fim da tubagem de admissão antes de mudar para uma solução tampão diferente. Lave a câmara de amostras com 100 uL neutravidina solução (ml em PBS / 0,5 mg) e incubar 15 min a temperatura ambiente. lavara solução neutravidina com 500 ul de PBS. Parafuso do suporte do metal para o prisma para a câmara de amostra. Montar a câmara da amostra para a fase micrómetro da TIRF-microscópio. Certifique-se de montar a câmara de amostra horizontalmente o mais reto possível na frente da objectiva para evitar desfocagem durante o processo de digitalização. Inicie o software que controla as câmaras EM-CCD e o software para controlar os motores piezoeléctricos do palco. Ajuste o foco da objetiva do microscópio, olhando para os reflexos do laser de IR. Coloque o prisma (PS991, N = 1,52) na parte superior do suporte de prisma de metal. Ajustar a posição lateral do prisma para certificar-se de que os feixes de laser atingiu o prisma em seguida, utilizar um adesivo e incubar com luz UV durante 5 min. NOTA: Prisma Montada pode ser reutilizado por limpeza. No "Aquisição Setup" software de controle de câmera clique e definir os seguintes parâmetros de aquisição: 100 ms INTEGRATtempo ion, 401 frames / filmes (câmera verde), 400 quadros / filme (câmera vermelho), multiplicadores de elétrons ganham 225, pré-amplificador de 5x de ganho e de leitura taxa de 3 MHz a 14 bit. Crie uma pasta no disco rígido local para a medição. Escolha um nome desejado para os arquivos de medição, por exemplo, ano-mês-dia. No software configurações de ir para o "Auto-Save" cavaleiro, permitir "Auto save" e escolher o formato de arquivo * .sif para a aquisição filme. Selecione a pasta no disco rígido. Use o nome da pasta como filestem. Ativar o "AutoIncrement" (valor ajustado início a 1) função. Permitir a ligação de um operador para o nome do arquivo. Use "DON" e "ACC" para o doador e do canal receptor, respectivamente. Escolha "_" como separador. No software de controlo da câmara, clique no "Video" para iniciar a imagem ao vivo da câmera e fundo lixívia de fluorescência por digitalizar a câmara de amostra usando intensidade máxima do laser de todos os três lasers (COMcombinada ≈3,000 W / cm 2 para 10 s por campo de visão). Desligue o laser azul. Diminuir a intensidade do laser verde a cerca de 200 W / cm2 e cerca de 40 mW / cm 2 para o laser vermelho alternadas se excitação com laser (Alex) é usado. Dilui-se a amostra fluorescente biotinilado até uma concentração de 50-100 pM. Carga de 100 mL da solução. A amostra está imobilizada na superfície da câmara após ligação. NOTA: Certifique-se de não sobrecarregar a câmara. moléculas vizinhas devem ser separados uns dos outros. Se necessário, carregue um adicional de 100 ul de uma amostra 2x mais concentrado para a câmara. Veda-se o tubo de entrada e de saída da câmara de medição, utilizando grampos após o carregamento é concluído. Desligue todos os lasers e usar os piezo-motores para mover a câmara de fluxo dois campos de visão ainda mais. No software de controle de câmera clique em "Tome sinal" para iniciar a gravação do filme e alternarsobre o laser ao mesmo tempo. Assegure-se que mais de 80% das moléculas são branqueadas até ao final do filme ajustando a potência do laser. Repita os passos de 3,17 e 3,18 para a região câmara de amostra pré-branqueado todo. 4. Aquisição do Mapa de Transformação ( "beadmap") Preparar uma câmara de fluxo conforme descrito nas secções 1.1, 1.2 e 2. Use esferas multi-espectrais fluorescentes revestidas com avidina, que mostram a emissão de fluorescência do dador e do aceitador de canal. Vortex a lingua durante 1 min, depois dilui-se 50 ul de estoque em 50 mL ddH 2 O. Vortex novamente durante 1 min, sonicar 1-2 min, depois vortex mais 10 s. Realizar passos descritos para medições smFRET (Seção 3,5-3,10). Carga de 100 uL (1) o volume da câmara de as 1: 2 diluídas esferas fluorescentes para dentro da câmara de fluxo. Esperar 10 min para as pérolas fluorescentes para se ligar à superfície. Use os parâmetros de aquisição em 3,9, mas change duração do filme a 26 (câmera verde) e 25 (câmera vermelho) eo ganho de multiplicador de elétrons a 10. Definir a intensidade do laser verde para um valor de 20 W / cm2. Tome um filme em um campo de visão com aproximadamente 50-100 esferas. 5. Processamento e Análise de Dados smFRET Utilizar o costume escrito de FRET SM software para a análise da beadmap (ver Materiais e Métodos) e os filmes adquiridos. Inicie o programa viewPlot1.m. Clique em Análise | Análise Batch, desmarque a opção "Alex" se não tiver sido utilizado. Para um melhor desempenho escolher o limiar de pico descoberta "alta". Pressione OK". Escolha "Não" quando perguntado se um beadmap já foi analisado. Procure a pasta que contém o beadmap adquirida e selecione o arquivo .sif * (clicando duas vezes sobre ele). Na janela seguinte, prima diálogo "OK". NOTA: Se um beadmap já analisou em uma medida anteriormento, escolha "SIM" aqui e selecione o beadmap salvas, navegando até a pasta correta e clicando duas vezes no arquivo * .map beadmap. Continue com o passo 5.8. Escolha duas contas individuais posicionados em cantos opostos do campo de visão. As intensidades de pixel são-codificados por cores de azul escuro (baixa intensidade) ao vermelho escuro (alta intensidade). Clique no centro do primeiro cordão. Se o centro da molécula pode ser localizado de forma clara pela codificação de cores, escolha "SIM" ou então clique em "NÃO" e escolher um par molécula diferente. Posicionar o cursor sobre o pixel, mostrando a intensidade máxima e pressione "Keep". Repetir o processo com o segundo canal. Clique na molécula no canto oposto e repita os passos 5.5 e 5.6. NOTA: O deslocamento de pixels relativa dos dois canais é exibida na janela de comando eo mapa de transformação é automaticamente salvo como arquivo * .map para a pasta que contém o beadmap * .sif arquivo <./ Li> Para carregar os filmes doador e receptor (* .sif) para a "análise de lote", navegue até a pasta, selecione todos os filmes que serão analisados e clique em "OK". Na janela de diálogo seguinte, pressione "OK". NOTA: A análise lote termina quando a última faixa exibida na janela de comando começa com "análise acabamento …". As moléculas detectadas são exibidos em uma nova janela, o que também indica o desvio relativo do dador e aceitador do canal determinada a partir do mapa de transformação. Para carregar os arquivos de filme em lote clique em File | Load. Desmarque a opção "Alex" se não tiver sido utilizado. Defina o smoothwidth a 10 e clique em "OK". Escolha a pasta que contém os arquivos * .ttr e clique em "selecionar tudo" e "OK" na próxima menu de contexto. Se o traço exibida apresenta as fases características smFRET (Figura 2) Pressione o botão de alternância "Não selecionado" e primeiroselecionar o ponto de tempo do início do evento FRET movendo a linha com o cursor do mouse e clicando no botão esquerdo do mouse. Próximo seleccionar o ponto de tempo do branqueamento da molécula receptora e, finalmente, o ponto de tempo do branqueamento da molécula dadora. Na janela seguinte, a eficiência FRET é representada em azul. Para selecionar a imprensa traço no botão "Sim", caso contrário escolha "Não". Para voltar a aceder a um traço do tempo, clique no botão "Anterior". Repita o procedimento até a última molécula do filme. Depois de analisar a última molécula em um filme salvar os traços selecionados, clicando em "Arquivo | Salvar". Salvar os traços selecionados na mesma pasta que os arquivos * .sif. Repita os passos de 5,10-5,13 para todos os filmes adquiridos. Execute o combine_fret_results.m programa. Selecione a pasta que contém os arquivos * res e todos os arquivos * .FRETonly_trace. Salve os arquivos FRET FRET e quadro-wise molécula-wise como MW.dat eFRW.dat, respectivamente. NOTA: Os arquivos * .dat são salvos como arquivos ASCII. O arquivo FRW.dat contém seis colunas e uma linha para cada FRET-frame. A sexta coluna contém a eficiência FRET frame-sábio corrigido. O arquivo MW.dat contém 21 colunas e uma linha por molécula FRET selecionado. A terceira coluna contém a eficiência FRET molécula-wise. 6. Resultados smFRET dados em histogramas NOTA: Para extrair a eficiência smFRET média de todos os dados smFRET gravados os dados do quadro-wise ou os dados molécula-wise são plotados em histogramas e analisados usando Gaussian cabe (múltiplos) picos. No que se segue, o protocolo usa um software de análise de dados comercial (ver Lista de Materiais). No entanto, qualquer outro software disponível pode ser usado em substituição. Abra um software de análise de dados (veja a Lista de Materiais). Clique em File | Import | ASCII múltipla. Selecione a pasta que contém o arquivo FRW.dat. Selecione o arquivo e pressione "OK & #34 ;. Aceite a opção de entrada com "OK" sem uma mudança. Escolha a terceira coluna C (Y) contendo as eficiências FRET corrigidos, clique com o botão direito na coluna e selecione Plot | Estatísticas | Histograma. Na janela do histograma, clique duas vezes nas colunas e desmarque "binning automática" e selecione um escaninho de tamanho desejado, por exemplo, 0,05. Também escolher os valores inicial e final, por exemplo, -0,025 e 1,025. Selecione as colunas do histograma clicando com o botão esquerdo sobre eles. Em seguida, clique com o botão direito e escolha "ir para Bin Planilha". Selecione a coluna "conta" clicando com o botão esquerdo sobre ele e, em seguida, clique com o botão direito e selecione Plot | Coluna / Bar / Pie | Coluna. Na trama bar coluna ir para Análise | Montagem | Nonlinear ajuste de curva | diálogo Abrir. Escolha "Gaussian" em função, em seguida, ir para o piloto "Parâmetro". Desmarque auto parâmetro de inicialização. Corrigir o valor do deslocamento (y0) em 0. Clique em "Ajustar". NOTA: A função de ajuste, bem como o de montagemcaudas são agora apresentados na trama coluna. O valor "xc" indica o centro da função de ajuste, ou seja, a média da eficiência de FRET, que serve como o parâmetro de entrada para o software NPS. 7. A medição do rendimento quântico Executar a determinação do rendimento quântico com o método relativa semelhante à do procedimento descrito por Würth et ai. 35, usando rodamina 101 dissolvido em etanol (QY = 91,5%) como um padrão. Gravar os espectros de absorção a um espectrómetro de UV-VIS, utilizando um volume de 80 mL numa cuvete de absorção de comprimento de percurso de 1 cm. A absorvância no comprimento de onda que vai ser usada para excitação de fluorescência tem de ser igual ou inferior a 0,05. espectros de emissão registro em um espectrômetro de calibrado-lamp operado em modo de fóton-contagem. Realizar as medições com polarizadores Glan-Thompson na excitação (0 °) e emissão (54,7 °) caminho (condições ângulo mágico), utilizando um spectr ai largura de banda de cerca de 5 nm e 2,5 nm para a excitação e emissão monocromador, respectivamente. Medir amostras depois transferi-los para uma tina de fluorescência com 3 milímetros de comprimento caminho cuidar que a taxa de contagem não exceda 10 6 s -1. Calcular o rendimento quântico de acordo com a em que n e n STD são os índices de refracção do solvente da amostra e o padrão, respectivamente. f (λ) e f_Std (λ) são as intensidades de fluorescência da amostra e do padrão no λ comprimento de onda. Um (λ ex) e um std (λ ex) são a absorvância da amostra de referência e no comprimento de onda de excitação e Φ STD é o rendimento quântico do padrão. le "> 8. Cálculo do Isotrópica Förster Radius Calcular o raio isotrópico Förster ( ) A partir do espectro de emissão da molécula dadora, o espectro de absorção da molécula receptora, o rendimento quântico do dador e o índice de refracção do meio. Use o PhotochemCAD freeware para o cálculo do . No entanto, qualquer outro software disponível pode ser usado em vez de 36. 9. A medição das anisotropias Determinar anisotropias de fluorescência em estado estacionário de gravações de espectros de fluorescência com várias configurações polarizador de excitação / emissão (v / v, V / H, H / V, H / H) 36. Calcular a-fator G, que corrige para artefactos polarização do instrumento, para cada comprimento de onda dorelação e usá-lo para calcular o valor de anisotropia para cada comprimento de onda: onde eu xy indica a intensidade de excitação polarização x e polarização emissão y. Calcular a média dos valores em toda a gama espectral de emissão de fluorescência para calcular a anisotropia do estado estacionário. 10. Instalação de Software Fast-NPS Baixar UCSF Chimera de http://www.cgl.ucsf.edu/chimera e seguir o guia de instalação. Ir para o site do "Instituto de Biofísica"em Ulm University: https://www.uni-ulm.de/en/nawi/institute-of-biophysics/software.html. Baixe a versão atual do Fast-NPS e extraí-lo para uma pasta de escolha. Abra a subpasta "Redistributable" e instale o ++ Redistributable Visual C, que é apropriado para o sistema. 11. Centrar o arquivo PDB Abra o arquivo pdb (s) de interesse em Chimera. Selecione todos os átomos do complexo macromolecular e calcular as coordenadas do centróide (Ferramentas | Análise de estrutura | Eixos / Planos / Centróides | Definir centróide … | Ok). Abra o Log Responder (Favoritos | Responder Log) e da Ferramenta de Transformação (Ferramentas | Movimento | transformar as coordenadas). Introduzir as coordenadas do centróide mostrado na resposta Log na caixa de texto "Shift" da janela de transformar as coordenadas e mudar o sinal de todas as coordenadas. Pressione o botão "Aplicar" e salve o arquivo com "Save PDB" (Arquivo | Salvar PDB). 12. Definiçãoos priores Posição NOTA: Todos os valores são considerados na angstrom. Inicie a linguagem de computação técnica e alterar a pasta actual para a pasta Fast-NPS local. Digite na janela de comando: FastNPS. Criar um novo jobfile no Project Manager (Project | New). Configurar a posição anterior (Ferramentas | corante modelo anterior). No painel "fundamentos anteriores" definir a resolução espacial da posição anterior, digitando seu valor (2 é recomendado). Excluir o interior da macromolécula, ativando a caixa de seleção e clicar no botão "load PDB". Selecionar e carregar o arquivo PDB centrado conforme descrito na Seção 11. Especifique o diâmetro aproximado (13 Å é recomendado, ver discussão) do corante, digitando o seu valor. Introduzir uma distância esqueletonização, ou seja, a distância que a molécula de corante pode penetrar na macromolécula (2 A é recomendado). Nopainel de "tamanho máximo antes" introduzir as coordenadas mínimas e máximas da posição anterior (recomendado: x em [-150.150], y em [-150.150] e z em [-150.150]). Ao definir um satélite, ative a opção "anexo via ligante flexível" no painel "fundamentos anteriores" e entrar no painel "ligador" as coordenadas do átomo (no arquivo pdb centrado) na qual a molécula do corante é anexado. Além disso, especificar o comprimento e o diâmetro do ligante ao introduzir os valores (13-A e 4,5 A são recomendados, ver discussão). Em caso de uma antena ignorar este ponto. Pressione o botão "calcular o volume acessível". Salvar a posição anterior e, opcionalmente, exportá-lo para fins de visualização utilizando o software como Chimera. 13. Definir a geometria da rede Abra a janela Definir Medição (Modo | Edit Geometry). Criar uma nova molécula do corante pressionando o bumbumem "New" no "Corantes" do painel. Defina sua anisotropia de fluorescência (Seção 9) inserindo um valor e selecione um modelo de corante dentro do menu suspenso "modelo Dye". Pressione o botão "Load", selecione a posição correspondente antes e marque a caixa de seleção Ativar do corante. Repetir este procedimento para todas as tintas, isto é, para todas as antenas, bem como para todos os satélites. Depois de criar todos os corantes, definir as medições. Criar uma nova medição, clicando em "New" no "Medições" do painel. Selecione seus parceiros FRET nos menus suspensos "Dye1" e "Dye2" abaixo. Digite a eficiência smFRET com o erro e do raio Förster isotrópico deste par corante. Por fim, marque a caixa de seleção Ativar da medição. Repita este procedimento para todas as medições. NOTA: Muitas vezes a rede se torna cada vez mais complexo, de modo que o usuário pode ficar confuso. A fim de Prevent erros, verifique a rede visualmente, premindo o botão "Verificar rede". A figura mostra as tinturas activados e indica as medições por meio de linhas que interligam os corantes FRET. 14. Cálculo Abra a janela de cálculo (Modo | Cálculo). Se cada corante na rede tem um modelo específico atribuído, selecione "Definido pelo usuário" e iniciar o cálculo pressionando "Cálculo". Para ter todos os corantes no mesmo modelo, selecione um dos cinco modelos (clássico, iso, meanpos-iso, var-meanpos-iso e var-meanpos) e continuar. NOTA: A janela de comando irá indicar o progresso do cálculo. Fast-NPS vai fazê-lo com uma mensagem pop-up, quando o cálculo tiver sido concluída. 15. Visualização de resultados A fim de exportar os volumes credíveis dos corantes, abra a visualização Janela de Resultados (Modelo | Ver Resultados). densidades de corante de exportação: Exportarcorantes isoladamente ou todos ao mesmo tempo. Para exportar um único corante selecione-o no painel "Corantes exibida" e pressione "Densidade Export". Digite uma resolução (2 é recomendado) e escolha um tipo de arquivo para exportação. À direita a densidade é visualizado e algumas de suas características matemáticas são mostradas. Para exportar todos os corantes simultaneamente empurrar "Batch Export". Abra os arquivos de densidade, resultando em quimera. 16. Verificação de consistência de Chosen Modelo Combinação Abra a visualização Janela de Resultados (Modelo | Ver Resultados). Se no painel "Informações de cálculo", a caixa de texto "Consistência" apresenta um valor inferior a 90% o modelo atual não representa as eficiências smFRET medidos suficiente e é, portanto, inconsistente. No caso de inconsistência apertar o botão "Consistência detalhada". Procurar as medições que têm um valor abaixo de 90%. Se um ou mais de corantes são predominantemente envolvidos nestas medições, seus modelos são susceptíveis de causar a inconsistência. Considerar diferentes modelos de corante para estes corantes e execute novamente o cálculo Fast-NPS.
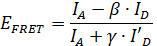 ,
, 

 ) Está definido para 2/3. Como resultado, os volumes são calculados credíveis quase duas ordens de grandeza menor, em comparação com aqueles no modelo clássico 32. No caso em que o fluoróforo é encontrada num ambiente que permite não só reori rápidoentação, mas o movimento adicionalmente rapidamente em todo o seu volume acessível, deve ser utilizado o modelo meanpos-iso. Neste modelo, o corante ocupa efetivamente apenas uma posição média, onde a média espacial é representada por uma conversão de distância polinomial 15. Este modelo aplica-se, por exemplo, a (geralmente hidrofóbico) corante está ligado a uma região hidrofílica, por exemplo, o ADN. Aplicação do modelo meanpos-iso leva a uma redução adicional do tamanho dos volumes credíveis por um factor de aproximadamente dois. No entanto, um corante ligado a uma proteína pode ligar-se reversivelmente a várias manchas hidrofóbicas em seu volume estericamente acessíveis (AV). Um fluoróforo que instantaneamente alterna entre essas regiões, mas dentro de uma região sofre livre rotação e movimento rápido localizado é melhor descrito pelo modelo VAR-meanpos-iso. Para uma situação semelhante em que o corante não é livre para girar VAR-meanpos modelo se aplica. Mais d etails sobre esses modelos podem ser encontrados em nossa recente publicação 32.
) Está definido para 2/3. Como resultado, os volumes são calculados credíveis quase duas ordens de grandeza menor, em comparação com aqueles no modelo clássico 32. No caso em que o fluoróforo é encontrada num ambiente que permite não só reori rápidoentação, mas o movimento adicionalmente rapidamente em todo o seu volume acessível, deve ser utilizado o modelo meanpos-iso. Neste modelo, o corante ocupa efetivamente apenas uma posição média, onde a média espacial é representada por uma conversão de distância polinomial 15. Este modelo aplica-se, por exemplo, a (geralmente hidrofóbico) corante está ligado a uma região hidrofílica, por exemplo, o ADN. Aplicação do modelo meanpos-iso leva a uma redução adicional do tamanho dos volumes credíveis por um factor de aproximadamente dois. No entanto, um corante ligado a uma proteína pode ligar-se reversivelmente a várias manchas hidrofóbicas em seu volume estericamente acessíveis (AV). Um fluoróforo que instantaneamente alterna entre essas regiões, mas dentro de uma região sofre livre rotação e movimento rápido localizado é melhor descrito pelo modelo VAR-meanpos-iso. Para uma situação semelhante em que o corante não é livre para girar VAR-meanpos modelo se aplica. Mais d etails sobre esses modelos podem ser encontrados em nossa recente publicação 32. 