All experiments were carried out in accordance with the accepted standards of humane animal care under the regulation of the IACUC at QU; animals were held in the zebrafish facility under Qatar University Biomedical Research Center (QU-BRC). All animals used in these experimental studies were under 3 days post-fertilization (dpf).
NOTE: For each experimental group, it is advisable to use at least 30 embryos for statistical rigor. The experimental groups are as follows:
Control group: This group includes embryos cultured in egg water without any injections. Results here will form the control baseline.
Negative Control group: This group includes embryos cultured in egg water injected with scrambled MOs.
Injected group: This group includes embryos injected with hand2 MO alone and hand2 MO with hand2 mRNA to rescue the phenotype. Results here will confirm that observed phenotypes appeared due to injected MOs. Comparison of experimental groups will enable assessment of the influence of inhibiting and rescue of hand2 on heart function precisely.
1. Morpholino designs for hand2.
NOTE: MO sequences can be adapted from the literature9,10,11. Alternatively, these oligos can be designed online by Gene-tools. Gene-tools offers a free and fast online design service, which can be accessed through their website12. A custom MO can readily be designed by providing information about the genes of interest, such as sequence information or accession numbers. The following specific steps summarize how to design MOs against hand2 in zebrafish:
- First, search for details of the gene of interest from the NCBI database. For hand2, in zebrafish13.
- Get GenBank mRNA transcript ID from NCBI14.
- Obtain the mRNA sequence from NCBI15.
NOTE: Morpholino (MO)-modified antisense oligonucleotides for the zebrafish hand2 gene are available and were previously published. hand2 MO sequence9,10,11: 5'-CCTCCAACTAAACTCATGGCGACAG-3', MO sequences used in the study are listed in the Table of Materials. - Get a negative control from either a mismatched or scrambled MO (similar sequences with random base pair changes) from gene tools16 to be injected. This would help attest to the specificity of the phenotype (s) observed in specific MO injections and minimize the risk that the observed effects are from an artifact of the injection procedure.
2. Preparation of morpholino injection
- Stock preparation
- Dissolve MO stocks with ddH2O; Diethyl pyrocarbonate (DEPC) can damage MOs and be toxic to embryos. Hence, use DEPC-free ddH2O. Upon ordering, MOs are delivered in a vial at a concentration of 300 nM.
NOTE: While Gene Tools recommend 1 mM stock solutions (~8 ng/nL), this can be too low, especially if the morpholino requires a high dose or is mixed with other MOs. Therefore, it is ideal for making various concentrations of MO stock solutions (2 or 3 mM). - Prepare a 0.2 mM working solution by sub-diluting the working solution (i.e., for 2 mM stock, 1:10 dilution). Prepare 10 µL of working solution by adding Danieaux's solution (7 µL), stock morpholino (1 µL), and of phenol red (2 µL from 10x stock). Phenol red gives the solution a dark red color to help trace the injected material in the embryo.
- Keep MOs at room temperature (RT) in airtight containers to prevent evaporation. Do not keep MOs on ice on the benchtop, as the solution may precipitate. RT handling is most appropriate.
NOTE: Gene Tools recommends either of two storage methods for MOs: The working solution can be kept at -20 °C (for many months to years) or RT in a sealed tube for long-term storage. If kept at -20 °C, heat the oligonucleotide solution at 65 °C for 10 min and vortex to completely dissolve the morpholino before use (no need to warm it up if kept at RT).
- Dissolve MO stocks with ddH2O; Diethyl pyrocarbonate (DEPC) can damage MOs and be toxic to embryos. Hence, use DEPC-free ddH2O. Upon ordering, MOs are delivered in a vial at a concentration of 300 nM.
- In vitro transcription of mRNA
NOTE: The in vitro transcription was used to generate mRNA of Hand2 from Plasmid HAND2 (NM_021973) Human Tagged ORF Clone to be microinjected into zebrafish embryos for rescuing the phenotype due to knocking down zebrafish hand2 using morpholinos, plasmid details used in the study are listed in the Table of Materials.- Linearize the plasmid DNA by an appropriate restriction enzyme and purify using a DNA purification kit (Table of Materials). Use the purified linearized DNA as a template for in vitro transcription to generate mRNA for injection as per the manufacturer's instructions (Table of Materials).
- Finally, inject 250 pg of human mRNA for the gene of interest to perform the rescue of phenotype to test whether the lack of a phenotypeis indeed from the loss of zebrafish gene by MO.
- Preparing to inject
- Pull a needle using a micropipettepuller (Table of Materials), following steps 2.3.2-2.3.6.
- Use 1 mm capillary tubes with filaments and a micropipette Puller. Turn the machine on and open the lid
- Turn the mode selection knob to NO.2 Heater. Using NO.2 heater, adjust the knob to bring the heat to 68 °C
- Turn the mode selection knob back to Step 1 position. Use four weights (2 Type Light / 2 Type Heavy)
- Place the capillary on the holder and close the lid. Press the Start button to get the pulled needle.
NOTE: This will generate capillary needles with a long taper - Cut the tip of the needles briefly, using forceps. Sharpen the tip of the pulled needle for easy penetration into zebrafish embryos using a capillary beveler for about 30 s, resulting in tips with diameters of about 15-25 µm.
- Calibrate the needle to estimate the amount of injection following steps 2.3.8-2.3.13.
- Briefly mix mRNA/morpholino solution before loading it into the needle to dissolve the particles that could clog the needle fully.
- Add ~3-4 µL of MO solution to the back part of the injection needle using a micron-tipped pipet tip. The injection needles are equipped with a small trench to facilitate capillary backfilling.
- Confirm that there are no bubbles in the injection needle. Mount the needle to the microinjector.
- Check that the tip of the needle is not clogged and that MO solution can flow out of the needle by pressing the injection pedal.
- Calibrate the volume of injection by measuring the droplet size. Put the tip of the needle in mineral oil on a micrometer slide. Press the injection pedal and adjust the injection (Eject) pressure (psi) and/or injection time (ms) until the ejected droplet diameter is 0.1 mm, corresponding to 0.5 nL.
- Ensure that the injection volume is around 0.5-1 nL. Adjust the volume by changing the injection (Eject) pressure (psi) or time (ms).
- For picoliter injector setup protocol, follow steps 2.3.15-2.3.16
- Use picoliter injector for zebrafish injections. Turn on the injector's feed pump.
- Prepare the injector according to the requirements shown below:
- P-balance: Ensure that P-balance is around 0. Ensure that it is slightly negative to prevent yolk flow back into the needle, diluting the MO solution in the needle. Conversely, if the backpressure is too high, the MO will constantly flow out of the needle even without the exertion of pressure on the pedal, resulting in variability and inconsistencies in the injected dose and the observed phenotypes.
- P-inject: 20-25 psi is ideal; change this to adjust injection volume. This can range from 10-30 psi but start with ~20 psi to check whether it delivers the desired volume.
- Injection time: Ensure to reduce the injection time to 300 ms. Prior to adjusting the injection volume, make adjustments to the time of injection.
- For morpholino injection, follow steps 2.3.18-2.3.19.
- Prepare the injection chamber as follows: Make 2% agarose from 0.6 g of agarose and 30 mL of E3 medium. Pour the agarose into a Petri dish and place an injection mold on the agarose using a TU-1 injection mold. Once cooled in a freezer, furrows will form for placing and stabilizing embryos (Figure 1A).
NOTE: The 60x E3M stock solution recipe consists of 5.0 mM NaCl, 0.17 mM KCl, 0.16 mM MgSO4·7H2O, 0.4 mM CaCl2·2H2O in 1 L ddH2O with a final pH of 7.6. - Transfer the collected embryos to the furrows using a transfer pipette, remove all excess E3 medium around embryos to prevent the floating of embryos (Figure 1B).
3. Injection of MO and mRNA solution into the yolk
- Insert the needle through the chorion, inject near the boundary of cell/yolk to the embryo side and press the injection pedal (Figure 1C).
NOTE: The injected MO solution/phenol red will not diffuse immediately within the yolk. A red spot will be observed in the yolk that gradually dissipates. The injected material should not exceed more than 10% of the embryo size. MOs can be microinjected into the yolk of newly-laid zebrafish eggs at up to the 8-cell stage because the MOs can easily be taken up in the cytoplasmic stream of the yolk sac by the developing cells17,18. In plasmid or capped mRNA injections, the injection should be performed into a single blastomere of a one-cell stage embryo. - Transfer the embryos into a Petri dish with E3 medium and incubate them at 28 °C until 3 dpf, with daily replenishment of E3 medium. At that stage, perform the phenotypic assessment using a microscope.
NOTE: MO solutions must be carefully diluted for each investigation to establish the lowest concentration needed to induce a specific phenotype. Embryonic lethality for each MOs must also be determined. At higher concentrations (above 4-10 ng, depending on the MO), MOs tend to elicit non-specific effects, such as brain or general cell death. - Co-inject MO with 250 pg of mRNA of human hand2 in the same way to confirm the rescue of phenotype induced by loss of zebrafish hand2 gene by MO.
4. Western blot to verify the success of morpholino knockdown
- Collect the embryos at specific time points (48 hpf, 72 hpf) and dechorionate if needed using the Pronase enzyme.
- Deyolk the embryos to remove Vitellogenin using a previously known methodology19.
NOTE: Vitellogenin is a phospholipo-glycoprotein that serves as a source of nutrients for the growing embryo. High-resolution 2D gel electrophoresis and vastly improved western blotting are made possible by deyolking the embryos and removing Vitellogenin. - Perform western blotting as described previously20.
5. Cardiac structure and function assessment:
- Live imaging of zebrafish: Imaging the ventricle
NOTE: Several heart function/hemodynamics parameters can be calculated by visualizing the beating heart of zebrafish embryo21,22. These parameters include cardiac output (CO), ejection fraction (EF), stroke volume (SV), fractional shortening (FS), and fractional area change (FAC) (please see previous papers involving detailed protocols for cardiac function assessment for zebrafish embryos22,23). The following steps briefly explain how to calculate those parameters for zebrafish embryos at ~3 dpf, a stage when the skin of these organisms is transparent, enabling visualization with bright-field microscopy.- Put a drop of 3% methylcellulose solution (RT) in the concave well imaging slide.
- Position the Zebrafish embryo in the well using a suitable plastic dropper (Figure 2A, B)
NOTE: Overfilling the well may cause fish displacement out of the well. - Gently mix the 3% methylcellulose drop with the E3 medium to stabilize the hatched embryo.
NOTE: To prepare 3% methylcellulose solution, dissolve 3 g of methylcellulose powder in 100 mL of PBS, or other mounting media, in a flask. Place a stirring magnet in the mixture's flask and place the flask on a magnetic stirring plate. Set the speed to "low" and keep it at 4 °C for ~1 day to dissolve all the clumps. Once the methylcellulose is completely dissolved, aliquot into small tubes and store at -20 °C. - Position the fish on its left, with its right side facing up and anterior point to the left to facilitate unambiguous imaging for the ventricle (Figure 2C).
- Under the microscope, zoom in on the embryo heart with 100x magnification and start recording for ~5 s. Ensure that the ventricle borders are inside the imaging window (Figure 2D).
- Record time-lapse movies using a high-speed camera and stereomicroscope at about 100 frames per second (fps) of the whole embryo (Figure 2C), beating ventricle (Figure 2D), and moving red blood cells (RBC) in major vessels, such as the dorsal aorta or the Posterior Cardinal Vein (Figure 2E), for cardiac function analysis.
- Save the movie in either AVI movie format or TIFF (or JPEG) image sequences format.
- Calculate the heart rate
- Calculate the time needed to record two consecutive frames. For 100 fps the time interval is equal to 0.01 s.
- Choose a known point from any recorded cardiac cycle (i.e., end-diastole or end-systole). Calculate the number of frames needed to repeat the cycle.
- Multiply the time interval from step 5.2.1 with the number of frames from step 5.2.2. The result is the time duration (in seconds) for one heartbeat.
- To calculate beats per minute, divide 60 by the number obtained in the previous step. For normal 3 dpf embryos, this should be around 150 bpm (2.5 Hz).
NOTE: Many software applications can calculate heart rate automatically from beating heart recordings such as ViewPoint24 and DanioVision25. - Analyze the cardiac structure using a record time-lapse movie at about 100 fps to check the whole heart to detect the presence of cardiac edema or any other structural defect.
NOTE: Figure 3A shows a normal 3 dpf embryo heart, and Figure 3B shows a 3 dpf heart with cardiac edema and heart looping defect (elongated ventricle).
- Calculate the fractional area change (FAC):
NOTE: Fractional area change (FAC) is a parameter used to compare ventricular end-diastole and end-systole areas to assess contractility of the ventricle.- Use a record time-lapse movie at about 100 fps to determine frames that represent one cardiac cycle. Extract the frames that show the end of both end-systole and end-diastole.
- Calculate both end-diastole area (EDA) and end-systole area (ESA) using ImageJ or a similar image analysis software.
- Use the following formula to calculate FAC22,26,27:
FAC
- Calculate the fractional shortening (FS):
NOTE: Fractional shortening (FS) is another parameter used to evaluate the contractility of the ventricle; to determine FS, the end-diastolic and end-systolic diameters of the ventricles must be measured21.- Repeat step 5.3.1
- Use ImageJ or any other equivalent image analysis program to measure the diameters of the ventricular walls at the end-diastole Dd and the end-systole Ds points. In most cases, short-axis diameters are employed to determine the fractional smoothness (FS) (Figure 4).
- Use the following formula to calculate FS22,27:
FS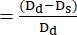
- Calculation of stroke volume (SV):
NOTE: For each heartbeat, the SV is the amount of blood pumped from the ventricle, which is easily computed from the end-diastole and end-systole volumes of the ventricles28.- Repeat step 5.3.1 and 5.3.2
- Measure the End-diastole (DL) and End-systole (DS) diameters as shown in Figure 4.
NOTE: Assuming that the ventricles of zebrafish hearts have a shape of a prolate spheroidal21,22, ventricle volume is calculated using the following formula.
Volume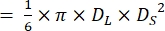
SV can be calculated following the below formula27. The normal SV range is 0.15-0.3 nL for 2-6 dpf embryos29. Here, EDV is end-diastole volume, and ESV is end-systole volume.
SV = (EDV – ESV)
- Calculate the ejection fraction (EF):
NOTE: EF is defined as the fraction of blood ejected from the ventricle with each heartbeat28.- From the above formulas, extract EDV and ESV for the fish.
- Calculate EF as follow22,27:
EF (%)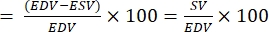
- Calculate the cardiac output (CO):
NOTE: CO is the volume of blood being pumped by the heart28. CO has a value of 10-55 nL/min for 2- 6 dpf embryos29.- Calculate SV and HR as mentioned in the previous subsections 5.5 and 5.2.
- Use the following formula to calculate CO22,27:
CO (nL/min) = SV (nL/beat) x HR (beats/min).
- Measure the RBCs velocities:
NOTE: Cells speed determination is vital for evaluating flow rate, vessel diameter, and calculating the shear stress exerted on the vessel's endothelial cells (i.e., Shear stress, on the other hand, is the frictional force imposed on the endothelial cells by moving blood). For 2-5 dpf embryos, the average red blood cells (RBC) velocity is about 300-750 µm/s29. To measure the RBCs velocities:- Under the microscope, zoom in on the animal tail at a magnification of 100x. RBCs movement should be visible at this magnification (Figure 5A, B).
- Start recording for about 8 s. Ensure that dorsal aorta (DA) and posterior cardinal vein (PCV), the two most important axial arteries, are 100-120 fps inside the imaging window. Track individual cells from sequential frames (Figure 5A, B).
- Extract several frames using ImageJ or other similar image analysis software.
- Calculate the difference in distance that an individual RBC moves (Δx).
- Determine the difference in time between the consecutive frames (Δt).
- Use the following formula for RBC velocity22,27:
RBC velocity (µm/s)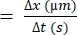
NOTE: Maximum and average velocity can be extracted from the repetition of consecutive frames for individual cells21. RBCs velocity also represents blood flow velocity. Alternatively, a variety of commercially available software applications can be used to measure RBCs' velocity automatically. ViewPoint24, as well as DanioVision25, have such applications (Figure 5C). Furthermore, there are few available plugins with ImageJ, such as TrackMate30 and MTrackJ31, compatible with movies recorded above 100 fps.
- Calculation of shear stress from measured cell velocities
Measured RBC velocities in blood vessels also represent blood flow velocities (Figure 5C). From these measurements, calculate the shear stress τ in a vessel of interest as follows with Poiseuille Flow assumption22,27:
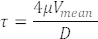
Where V is the average blood velocity (µm/s), µ is the blood viscosity (dynes/cm2), and D is the vessel diameter (µm). For 3 dpf embryos, shear stress in DA22,26,27 is about 4 dynes/cm2.
The graph in Figure 6 illustrates the average percent of embryos surviving at 24, 48, and 72 hpf for both HAND2- specific MO and control scrambled MO-injected embryos. The 1 mM (8 ng/µL) and 0.8 mM (6.4 ng/µL) MO-injected embryos showed a significant reduction in survival percentage compared to control scrambled MO-injected embryos. This was observed across each measured time point where lethality or malformation was observed. The results indicated that a high concentration of HAND2MO had a significantly lower survival percentage due to off-target effects and toxicity of MO. While 0.4 mM (3.2 ng/nL) HAND2 MO-injected embryos did not show a significant reduction in survival percentage; however, some similar phenotypes in the experimental group were noted, such as delayed in heart development, elongated tub-like structure heart, and pericardial edema in comparison to control scrambled MO-injected embryos (Figure 6).
HAND2 knockdown embryos exhibited distinct and specific phenotypes. Phenotypes of the morphants were examined between 24 and 72 hpf under a Light Microscope. HAND2 loss of function gave rise to embryos with a delay in heart development and an elongated heart, shaped like a tube with pericardial edema (Figure 7). As a result, embryos with this defect in heart formation behaved differently in terms of cardiac performance in comparison to scrambled MO (Figure 8).
In our investigation, we have characterized the changes in the zebrafish heart shape, cardiac output, and blood flow in control and HAND2 knockdown animals. Blood flow velocity was measured by tracking down moving RBCs, which were used to determine shear stress levels, an important mechanobiological factor on endothelial cells.
Cardiac output and blood flow analyses showed that HAND2 knockdown triggered a delay in heart development in comparison to wild-type embryos. This alteration resulted in a decrease in DA blood velocity, a drop in heart pulse, and a reduction in cardiac output (Figure 8A). In agreement with the negative impact of HAND2 knockdown on cardiac function, shear stress was also reduced significantly (Figure 8A) in comparison to the control scrambled MO-injected embryos. Most of the heart function failure was rescued when HAND2 MO was co-injected with human HAND2 mRNA, which resulted in an increase in DA blood velocity, increase in heart pulse, and restoration in cardiac output (Figure 8A). These results are in agreement with cardiac output and blood flow analysis results for PCV (Figure 8B). Collectively, the data suggested that HAND2 might play a critical role in cardiomyogenesis during zebrafish development.

Figure 1: Steps for injection of MO solution into the yolk. (A) Injection chamber. (B) Using a transfer pipette, transfer the collected embryos to the furrows. (C) Insert the needle through the chorion, inject near the boundary of cell/yolk to the embryo Please click here to view a larger version of this figure.

Figure 2: Mounting and imaging Zebrafish embryos. (A) The concave well imaging slide with zebrafish embryo under the microscope. (B) Magnified view of the well. Fill the wells ¾ of volume with E3 medium. (C) Positioning of the embryo should be on its left side. (D) Ventricle can be seen clearly in this configuration (zoomed image on left, ventricle borders are highlighted). Anterior is to the left. (E) Moving red blood cells (RBC) in major vessels, such as the dorsal aorta or the Posterior Cardinal Vein. Anterior is to the left. Please click here to view a larger version of this figure.
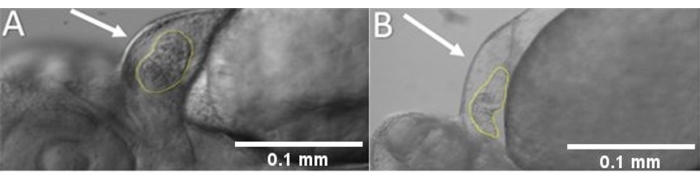
Figure 3: Analysis of the whole heart to detect the presence of cardiac edema and/or any structural defects. (A) A normal 3 dpf embryo heart. (B) Cardiac edema and looping defect in 3 dpf zebrafish embryo. Anterior is to the left. Please click here to view a larger version of this figure.
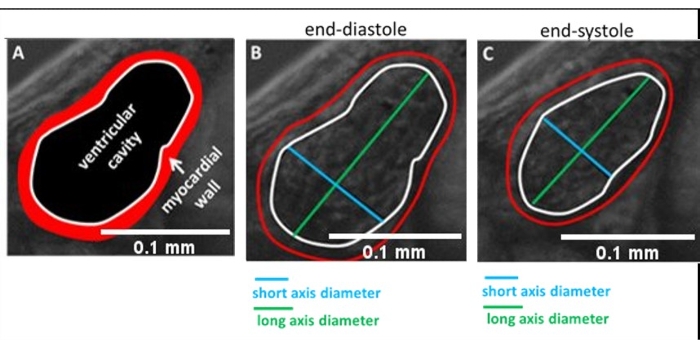
Figure 4: Measurement of ventricle size. (A) Ventricular cavity and myocardial wall are highlighted. Long axis and short axis diameters are seen for (B) end-diastole and (C) end-systole. Please click here to view a larger version of this figure.
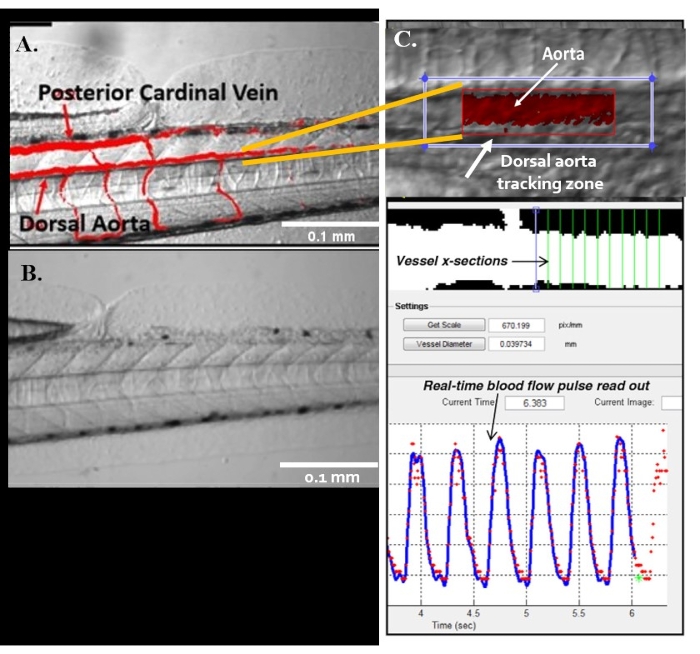
Figure 5: Automatic detection of vessels using ZebraLab. (A,B) The two major blood vessels in zebrafish: the dorsal aorta (DA) and the posterior cardinal vein (PCV). (C) Viewpoint program used to quantify blood flow and vessel diameter in DA. Please click here to view a larger version of this figure.

Figure 6: Hand2 MO titration. Embryonic survival for different Hand2 MO concentrations was converted to a percentage, averaged, and plotted. The percent survival of MO-injected embryos was calculated at 24, 48, and 72 hpf. All data points represent mean ± SEM (100 embryos were used in each group; experiment was performed in triplicate). The analysis was done by two-way-ANOVA with Dunnett test. *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001. Please click here to view a larger version of this figure.

Figure 7: Hand2 morphants exhibit distinct and specific phenotypes. Hearts and tails of 72 hpf embryos were examined under a light microscope (100x magnification). (A,B) Control scrambled MO shows normal heart development without pericardial edema. (C,D) 0.4 mM HAND2 MO-injected embryos as positive control shows tube-like heart structure. Please click here to view a larger version of this figure.
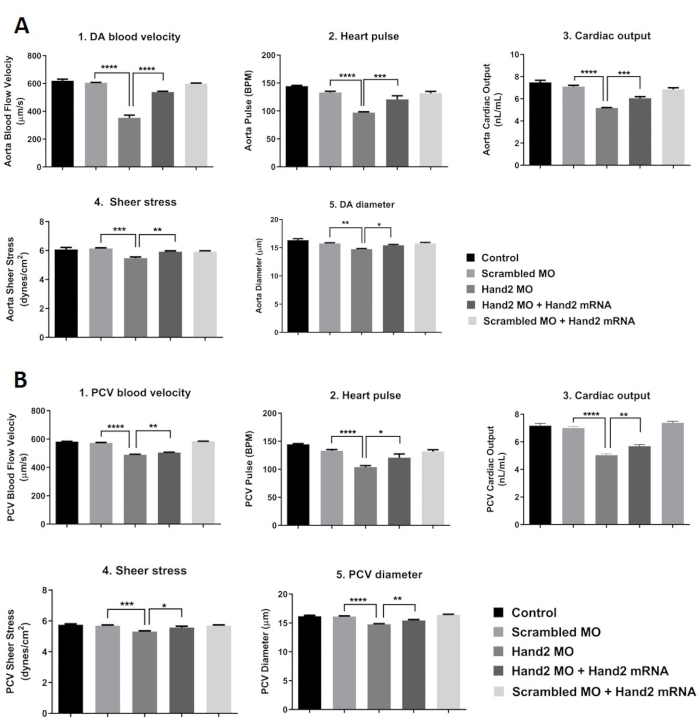
Figure 8: Assessment of cardiac function (A) Dorsal aorta blood flow analysis, (B) Posterior Cardinal Vein (PCV) blood flow analysis. 1-4 cell stage zebrafish embryos were injected as groups with scrambled MO, Hand2 MO, Hand2 MO + Hand2 mRNA rescue, and scrambled MO + Hand2 mRNA rescue. Un-injected embryos were used as control. All data are presented as mean ± SEM (6 embryos were used in each group; experiment was performed in triplicate). The analysis was done by one-way ANOVA with Sidak post hoc test. *p < 0.05, **p < 0.01, ***p < 0.001 and ****p < 0.0001. Please click here to view a larger version of this figure.
