The well-characterized PGPB P. simiae WCS417r is known to colonize the roots of A. thaliana in hydroponic culture. This naturally fluorescent bacterium can easily be visualized using microscopy on the roots of seedlings following colonization (Figure 2). Although it is possible to image the full length of these A. thaliana seedlings’ (4–6 mm length) roots, doing so for many plants would take a prohibitive amount of time. Because most variation across timepoints and species of bacteria can be captured by imaging the crown, middle, and tip of the root14 (indicated by red boxes in Figure 1), these regions were prioritized for imaging rather than imaging full root lengths. In the bright-field images of P. simiae-colonized A. thaliana roots (Figure 2), it is possible to visualize the outline of the roots and root hairs; however, at 18 h of colonization, it is not possible to clearly differentiate colonized versus non-colonized roots using bright-field images. While P. simiae displays autofluorescence, we used a strain also engineered to express a yellow fluorescent protein (YFP)19 with excitation/emission wavelengths of 490-510/520-550 nm18. A magnification of 100x was sufficient to clearly identify individual and small aggregates of P. simiae cells on A. thaliana roots. As shown in Figure 2, laboratories with access to either high-resolution confocal microscopes or less expensive benchtop microscopes can both visualize the presence and distribution of bacteria along the root.
While informative in terms of spatial distribution, microscopy images are not well-suited for quantification of bacterial cells. We thus collected bacteria from the surface of roots using ultrasonication as previously described and validated9,20. Three rounds of 12 s of ultrasonication21 at an amplitude of 40 were sufficient to disrupt the outer surface of the root seedlings (Supplemental Figure 1) and remove all bacteria while not impacting the bacterial viability. Sonication was used rather than bead-beating methods9 to better promote dispersal of bacterial aggregates/biofilms. Quantifying CFU/root after 18 h of colonization and an additional 72 h of maintenance showed that P. simiae both colonizes and is maintained on the roots of A. thaliana in our hydroponic, floating seedling system (Figure 3). The number of CFU/seedling at either timepoint showed good reproducibility across biological replicates performed on different days (Figure 3). The variation observed is common among root colonization assays22 and is likely due to minor variations of timing, environmental conditions, or plant root size, even among seedlings germinated at the same time and selected to be similar in size. We observe an increase in the number of CFU/seedling after 72 h in maintenance medium as compared to the numbers observed at the post-colonization 18 h timepoint (Figure 3). This indicates active growth of the colonized bacteria on the plant root occurred during the maintenance stage.
In addition to the utility of this hydroponic assay to quantify individual bacterial colonization and maintenance, it is also applicable to monitoring the association of multiple species on plant roots. To demonstrate this, three species of bacteria isolated from A. thaliana grown in natural soil under laboratory conditions were chosen20. The isolates were strains of Arthrobacter nicotinovorans, Microbacterium oleivorans, and Curtobacterium oceanosedimentum23. This simplified community was chosen due to these species’ ability to coexist in liquid bacterial growth media in shaking culture (unpublished data). In addition, these three species can be clearly differentiated on media containing X-gal due to differences in colony morphology and color (Figure 4A). The X-gal does not affect relative growth of any of these bacterial species (unpublished data). These differences in morphology and colony appearance on X-gal allowed us to count the CFU/seedling of each species without antibiotic selection, even in multi-species coculture.
A. nicotinovorans, M. oleivorans, and C. oceanosedimentum were all colonized and maintained on the root, whether alone or in bacterial coculture (Figure 4B). Each species showed trends that were similar across different biological and technical replicates, even within mixed communities. This demonstrates that the assay protocol can be used to measure both relative or total CFU/root of each species. Interestingly, when grown alone, no individual species showed an appreciable increase in abundance during the maintenance stage, but the overall CFU/root of the combined community increased in cocultures, indicating that these bacteria do not prohibit the colonization of the other strains.
For all experiments, plants grown in liquid media without the addition of bacteria as negative controls were always included. No bacteria were visible on these control roots during microscopy (Figure 2), nor were any bacteria detected via plating for CFU (unpublished data). This indicates that sterilization of seeds and using the sterile techniques during this assay were sufficient to keep plants axenic unless purposely colonized.
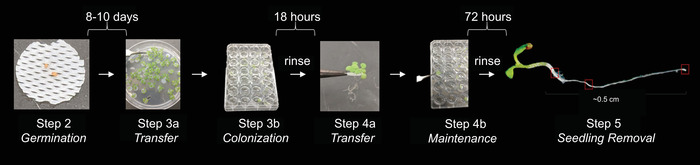
Figure 1: Assay for bacterial colonization and maintenance on A. thaliana roots. A. thaliana seedlings grown on sterilized plastic mesh were transferred to a growth medium optimized for bacteria [here, 0.1 x LB (Luria Broth) Lennox]. Bacteria then colonized the root over 18 h while the plant floated in shaking liquid. Following a rinse, the colonized float was transferred to a growth medium optimized for plants (0.5x MS + MES) for 72 h to test for maintenance of bacteria on the roots. The float was then rinsed, and the plant with any attached microbes was removed for analysis (quantification of CFU/seedling or imaging by microscopy). Please click here to view a larger version of this figure.
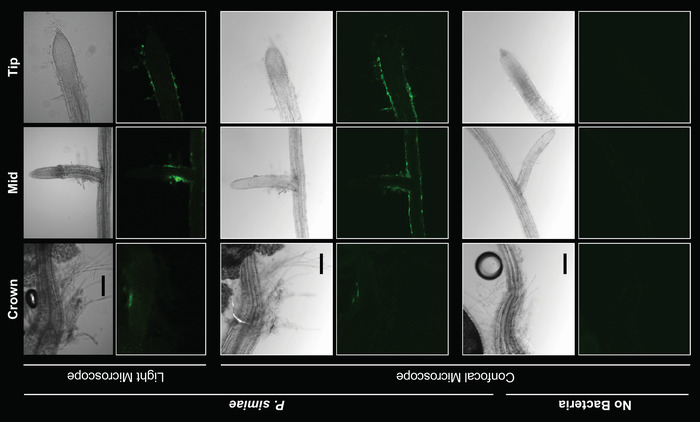
Figure 2: Visualizing P. simiae colonization of roots with fluorescent microscopy. P. simiae (false-colored green) colonized A. thaliana roots and was maintained on the root following transfer to plant-growth medium. Root crown (left), mid-length (center), and tip (right) at 40x magnification are shown from areas indicated in Figure 1. The top two rows show the bright-field and fluorescent images of roots colonized by P. simiae (imaged by epifluorescent microscopy). The same roots were also imaged by a confocal microscope (center two rows). The no-bacteria negative control in the two bottom rows showed no colonization. Scale bars represent 50 µm. Please click here to view a larger version of this figure.
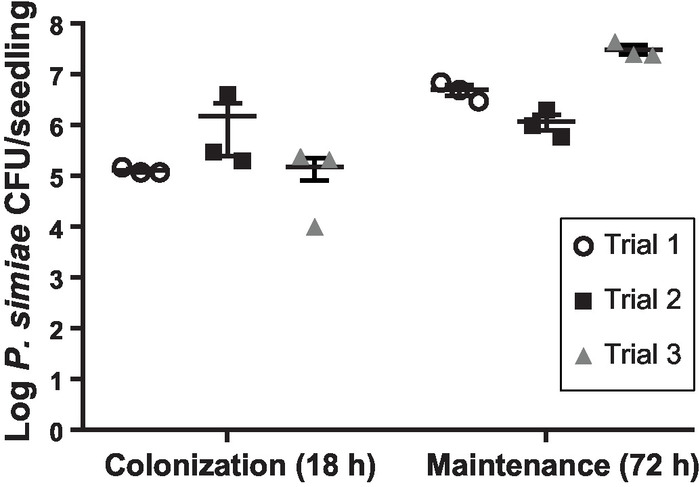
Figure 3: Quantification of P. simiae on A. thaliana roots. Total number of P. simiae viable cells recovered per A. thaliana seedling following 18 h of colonization or 72 h of maintenance. Three individual biological replicates are shown, each containing three technical replicates of two seedlings per float. The numbers shown are the means from the technical replicates, while bars represent standard errors of the means. Please click here to view a larger version of this figure.
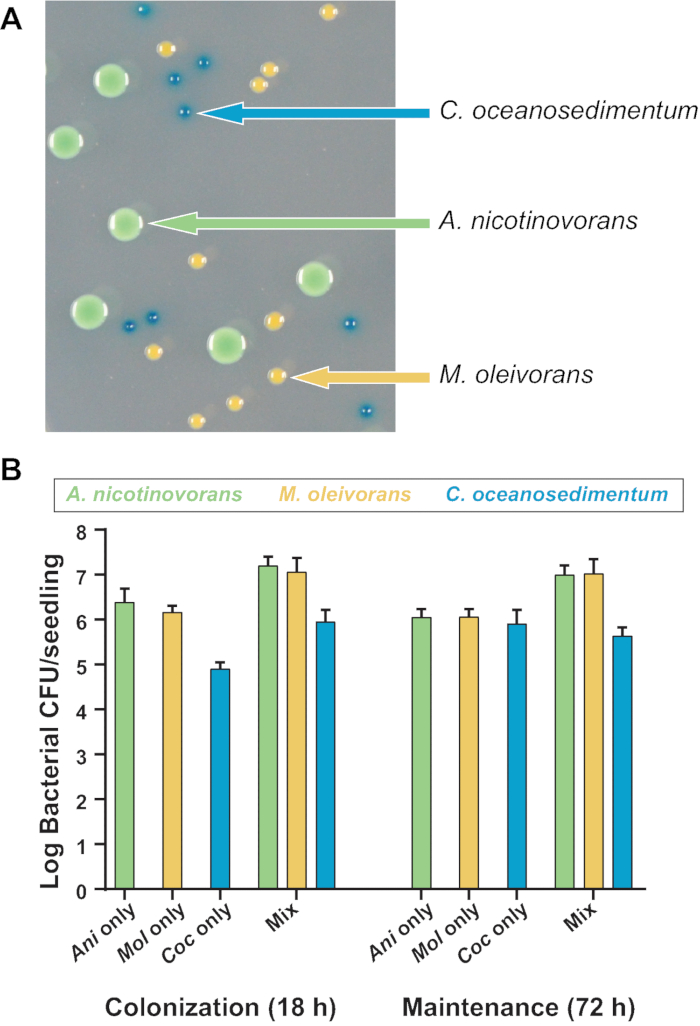
Figure 4: Quantification of the colonization and maintenance of a mixed bacterial community on A. thaliana roots. (A) Colonies of A. nicotinovorans, M. oleivorans, and C. oceanosedimentum can be differentiated on X-gal-containing agar medium by colony morphology and color. (B) Roots of 10 day-old seedlings were inoculated with approximately 3×105 CFU/mL of each of the three strains. Shown are total CFU/seedling recovered of each species following 18 h of colonization or 72 h of maintenance when colonized either alone or in a three-member bacterial community. Two biological replicates, each comprising two technical replicates of two seedlings per float, are shown. The numbers shown are the means from the two technical replicates, while bars represent standard errors of the means . Please click here to view a larger version of this figure.
Supplemental Figure 1: Ultrasonication disrupts the root surface. To dislodge bacteria from the surface of the root, whole plants were sonicated, and the bacteria was released into the liquid, which was serially diluted and plated for quantification of CFU/seedling. (A) An intact seedling is (B) structurally disrupted following ultrasonication. Please click here to download this file.
