Figure 1 shows stills of images taken from a video describing the dissection. Images provide typical locations of the nerves prior to cutting from the brain.
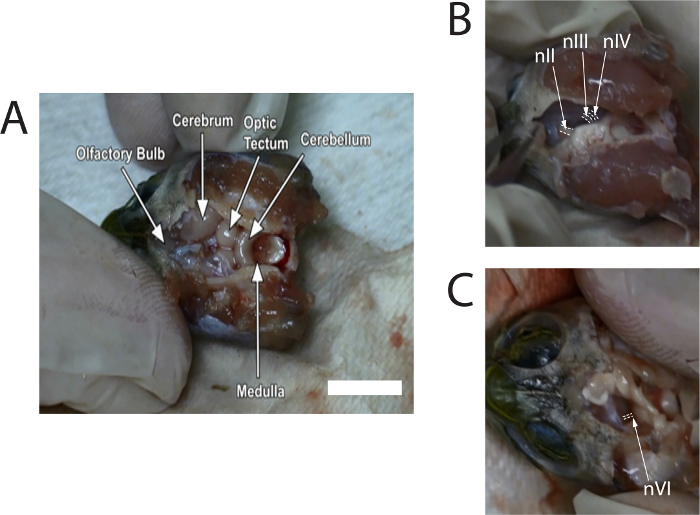
Figure 1: Stills of images captured from the video of the dissection to show locations of the optic nerve (nII), oculomotor nerve (nIII), trochlear nerve (nIV), and abducens nerve (nVI). (A) Image with labeled regions of the brain after removal of the meninx. White scale bar = 10 mm. (B) Image shows location of nII, nIII, and nIV where they connect to the right side of the midbrain (after removal of the olfactory bulbs and cerebrum). (C) Image shows location of nVI where it connects to the left side of the brainstem (after removal of the midbrain). White dashed lines are drawn around the nerves. Please click here to view a larger version of this figure.
Figure 2 shows the mean change in pupil diameter occurring after stimulating nIII. Although extrinsic eye movements also are observed, which change the location of the pupil, the measure of the action by the sphincter pupillae of the iris remains reliable. Measures are from one preparation and show the typical variability of responses to repeated current stimulations. Mean pupil diameter is reduced from 1.95 ± 0.01 mm to 1.60 ± 0.01 mm. The narrow standard deviation indicates a successful fit of the electrode to the nerve. When measuring among different preparations (N = 5), typical variability is ± 0.08 mm. Although still relatively narrow, the value is about eight times greater than the variability observed from a single preparation.
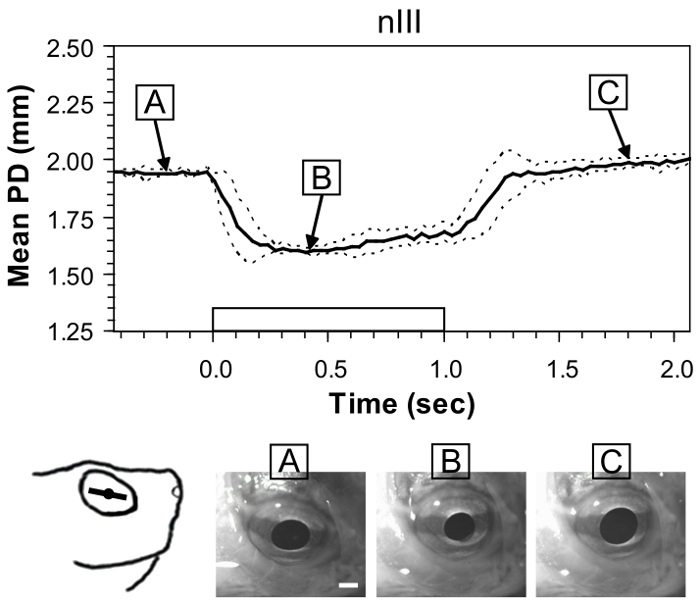
Figure 2: Example of pupil constriction evoked by stimulating the oculomotor nerve (nIII) in the whole-head preparation. Black trace is the mean pupil diameter (PD) from six stimulations in one preparation, and dashed lines show ± standard deviation (SD). Rectangular waveform on the x-axis denotes onset and offset of a 100-Hz train of 1-ms pulses with an amplitude of 50 µA. The sketch at the lower left shows the orientation ofthe iris line in the eye prior to stimulation,and images at the bottom show still frames from a representative trial before(A), during (B), and after stimulation (C). White scale bar = 1 mm. This figure has been reprinted with permission33. Please click here to view a larger version of this figure.
Measures of pupil responses do not require calibration of eye rotations; however, if measuring extrinsic movements, placement of the head in a gimbal must be carefully done to allow comparisons among preparations (Figure 3). When the head is placed in the gimbal with the dorsal surface of the skull parallel with the horizon, the iris line is offset from the horizon toward the nostril. Figure 3D shows a typical offset (28.6°) in a preparation. For three different preparations the mean offset was 30.1 ± 9.0°. An angle of offset within the standard deviation indicates acceptable fit within the gimbal.

Figure 3: An isolated turtle head preparation positioned in the gimbal. (A) Region of the white rectangle shows the location of the turtle head in a photograph of the equipment setup. (B) Magnified view of the white rectangle (seven times). Dotted white line is drawn on the image from nostril to pupil center and is level with the horizon. A solid white line is superimposed and parallel to the iris line. (C) Cartoon of the left side of the head. (D) Image of the eye captured by the camera of the video-based eye tracking system. Black scale bar: 5 mm in B; 1 mm in D. This figure has been modified with permission32. Please click here to view a larger version of this figure.
Figure 4 shows the mean rotations of the eyes from ten preparations evoked by stimulation of the nIV going to the superior oblique muscle. Typical peak rotations for intorsion, elevation, and abduction (in response to 70 µA, orange trace) are 10.0 ± 5.7°, 2.8 ± 1.2°, and 3.0 ± 2.1° respectively. Magnitudes measured for rotations in single preparations to repeated stimulations are similar but with less variability (e.g., N = 5), 14.8 ± 1.0° for intorsion, 3.2 ± 0.2° elevation, and 2.4 ± 0.2° for abduction. The pattern of variabilities among different preparations compared to a single preparation is comparable (within one logarithmic unit) to what is observed for measures of pupil changes: 6 times greater for both intorsion and elevation, 11 times greater for abduction.
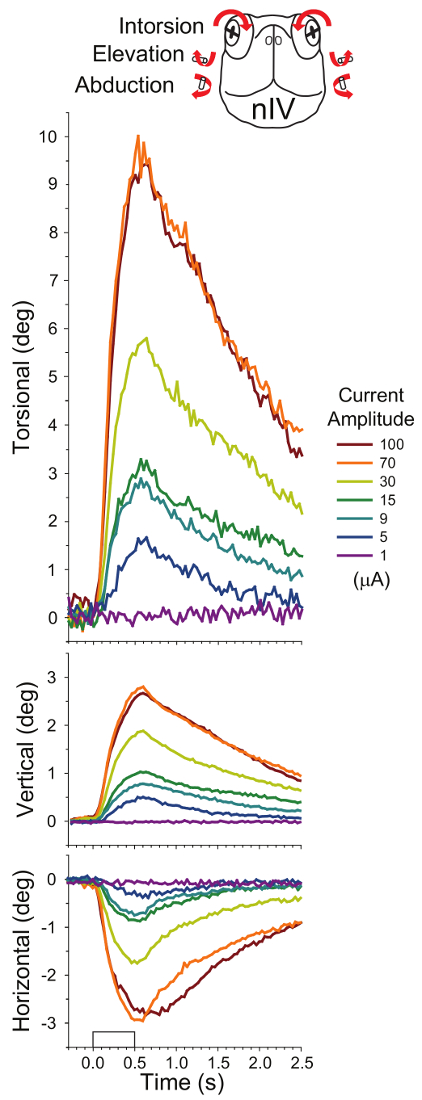
Figure 4: Mean eye rotations evoked after stimulating the left trochlear nerve (nIV) with 500-ms 100-Hz trains of 2-ms pulses in 10 preparations (six applied to the left side and four applied to the right side, five trials measured for each). The rectangular waveform on the x-axis of the bottom plot denotes timing of the stimulus. Responses from left and right eyes were not significantly different from each other. Intorsion was accompanied by elevation and abduction (see overlying cartoon of the head-on view of the turtle, whose arrows summarize the components of the eye movements). The eye movement amplitude roughly corresponds to the seven current amplitudes applied to nIV (see code in legend box). This figure has been reprinted with permission32. Please click here to view a larger version of this figure.
In contrast to nIV, which goes only to the superior oblique muscle, nIII and nVI innervate multiple muscles so movements are more challenging to interpret. Resultant eye movements depend on what motor units are recruited (Figure 5) (See Discussion). Consequently, variability from preparation to preparation can become significant. For example, nVI evokes intorsion, elevation, and abduction (Figure 5C). Abduction is likely from the motor units promoting the action of the lateral rectus; intorsion and elevation instead result from other motor units going to targets such as the retractor bulbi or the nictitating membrane.
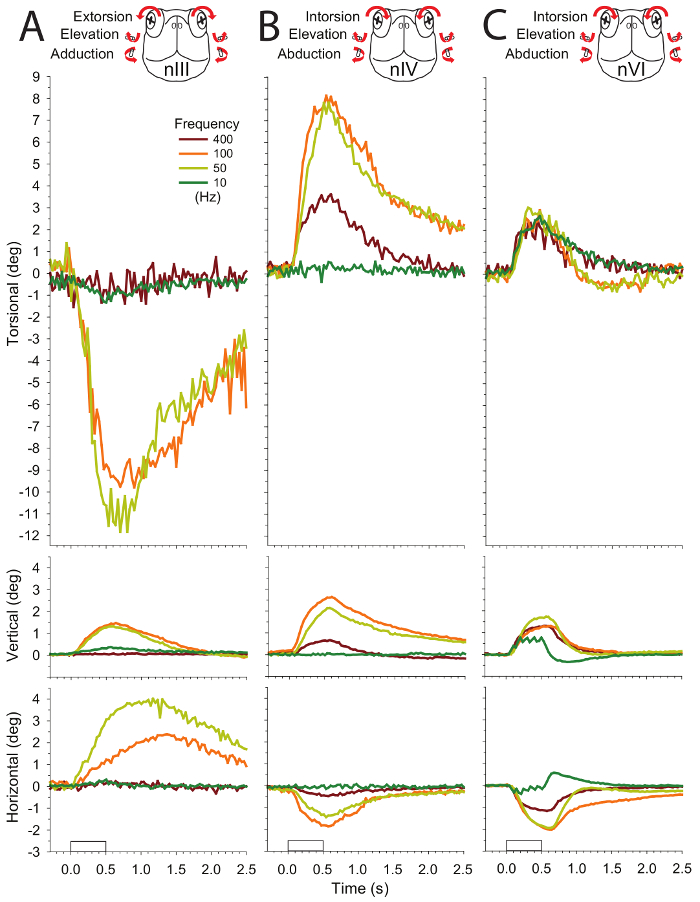
Figure 5: Mean eye movement responses evoked by stimulating three cranial nerves with 10-, 50-, 100-, and 400-Hz trains in four preparations. All of the stimulus trains were 500 ms in duration, consisting of 2-ms pulses with amplitude of 70 µA, a value above the threshold. Eye movements are displayed as before, with columns A–C showing responses to stimulation of nIII, nIV, and nVI, respectively. This figure has been reprinted with permission32. Please click here to view a larger version of this figure.
Because nIII innervates several targets (superior rectus, inferior rectus, inferior oblique, medial rectus, and the sphincter pupillae), analysis of the evoked movements are the most complex. For the example shown in Figure 5A, extorsion, adduction, and elevation occurs. Based on this, a reasonable interpretation is that the actions are mostly from the inferior oblique with possible contribution from the medial rectus. Superior rectus and inferior rectus may cancel each other. A higher variability of the actions for nIII likely stems from the greater branching of the nerve and the location of the cut for fitting the electrodes.
