Image acquisition of the glucose sensor in the ventral nerve cord (VNC), ex vivo
To determine differences in glucose uptake in a Drosophila model of ALS based on TDP-43, a genetically encoded FRET-based glucose sensor was used. The sensor comprised CFP and YFP fused to the glucose binding domain from the E. coli MglB gene. Glucose binding elicits a conformational change, which can be detected by fluorescence resonance energy transfer (FRET) between the CFP and YFP 10,11,12. When glucose is not bound to the sensor, CFP excitation causes CFP emission only, and when glucose binds, the YFP signal can be detected due to FRET occurring between CFP and YFP (Figure 1A). The D42 GAL4 driver was used to express the sensor alone or together with the ALS-associated mutant TDP-43G298S in motor neurons via the bipartite UAS-GAL4 expression system14. Third instar larvae were dissected in HL3 buffer without glucose and pinned to expose the neuromuscular system, including the VNC still connected to the neuromuscular junctions via motor neuron axons. The VNC was imaged using a 40x water immersion lens with a spatial resolution of 512 x 512 pixels (see Protocol and Figure 1B). CFP and FRET signals were acquired in the 465-499 nm and 535-695 nm, respectively, following CFP excitation with the 405 nm laser. The slice to be imaged was chosen based on the plane where the VNC was visible with a minimum of six motor neurons in focus along the midline. The selected plane was imaged for 10 min, capturing an image every 10 s to determine the baseline fluorescence. The czi files were saved as: genotype_sample#_baseline. After the 10 min time course was completed for baseline conditions, the buffer was replaced with glucose supplemented HL3. The image was refocused to find the same plane that was imaged under baseline conditions. The same image settings were used for all samples. The VNC was then imaged again for 10 min acquiring a new image every 10 s and saved as: genotype_sample#_post stimulation for further analyses.
Image analysis
A significant challenge with live imaging experiments is the drift that VNCs undergo during the 20 min imaging interval. To address this issue, a drift correction plugin, Stackreg was used in FIJI prior to image analysis and ROI selection (see Protocol and Figure 2A). 6-8 individual cells/ROIs were selected from each VNC and outlined using the FRET/YFP channel. Selections were copied to the CFP channel, and the mean gray values were measured using the multi-measure function. Quantifications were performed by first averaging the FRET/CFP ratios per time point, and then calculating the average for the entire 10 min per genotype. These averages were then used to normalize the FRET/CFP ratios at each time point and plotted over time to generate a first evaluation of the data (Figure 2B). Using this normalization approach, it was found that upon stimulation, control motor neurons expressing glucose sensor alone, exhibit a small but significant increase in glucose uptake (3.9%, Pvalue < 0.0001) while motor neurons expressing TDP-43G298S show a significantly higher glucose uptake (16.76%, Pvalue < 0.0001). These data are consistent with TDP-43 proteinopathy causing increased glucose uptake in motor neurons, as previously shown in reference9.
Data analyses – measurement of glucose uptake in motor neurons in the Drosophila VNC
To gain further insights into differences between controls and TDP-43G298S mutant motor neurons, each FRET/CFP ratio was normalized to the control at each time point (Figure 3A). This allows for a comparison in glucose uptake between ALS motor neurons expressing TDP-43G298S and control motor neurons expressing the glucose sensor alone in real-time. Under baseline conditions, there was no difference between the genotypes. However, upon stimulation with glucose, a rapid (7.31% after 1 min of glucose application) and significant increase (12.76% after 10 min, Pvalue < 0.0001) in glucose uptake was observed in TDP-43G298S when compared to the control.
Bar graphs offer an alternative approach to illustrating differences between genotypes (TDP-43G298S versus controls) before and after glucose stimulation (Figure 3B). This was done by normalizing the FRET/CFP ratio for both conditions to the baseline of the respective genotypes and plotting the average ratio for the last 5 min of baseline and stimulation imaging, respectively. Upon stimulation, neurons expressing glucose sensor alone show a small but significant increase in FRET/CFP ratio (3.83% ± 0.08%, Pvalue < 0.0001) compared to the baseline. Whereas neurons expressing TDP-43G298S showed a significant increase in FRET/CFP ratio upon stimulation (14.73% ± 0.08%, Pvalue < 0.0001) when compared to the baseline. The net difference in FRET/CFP ratios between motor neurons expressing TDP-43G298S and glucose sensor alone is 10.9% (Pvalue = 0.005). This analysis also shows that the ratios stabilize around the last 5 min of each time-course imaging, so this time frame was chosen for the graph. This analysis provides a high-level overview of the results, including statistically significant differences between genotypes and treatments in one graph.
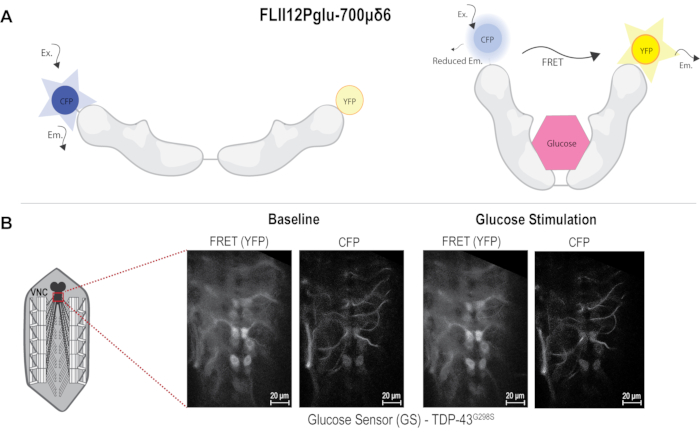
Figure 1: FRET sensor and image acquisition. (A) FRET-based Glucose sensor diagram used to measure glucose uptake in motor neurons. Glucose binding causes FRET to occur. (B) Diagram of neuromuscular junction (NMJ) dissection showing area imaged in the VNC. Images to the left showing TDP-43G298S before and after glucose stimulation in both FRET (YFP) and CFP channels. Scale bars: 20 µm. Magnification: 40x. Please click here to view a larger version of this figure.
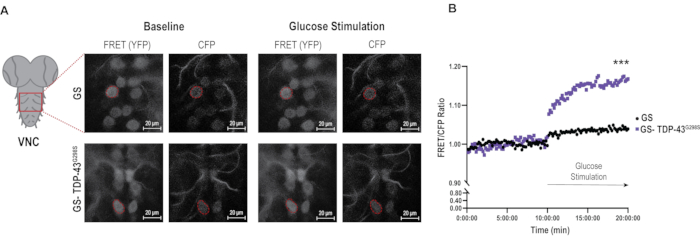
Figure 2: Image analysis of glucose sensor in the context of TDP-43 mutant. (A) Representative images of FRET and CFP signals in glucose sensor controls and TDP-43G298S samples under baseline and stimulation conditions. A representative motor neuron is outlined in each image as an example of an ROI. Scale bars: 20 µm. Magnification: 40x. (B) The average ratios of FRET/CFP in glucose sensor and TDP-43 mutant motor neurons over 20 min of imaging. Ratios were normalized to the average baseline ratio for each genotype (see baseline normalization in the Protocol). Values shown are the average of 25-30 motor neurons (ROI) from 5 VNCs per genotype. Mann-Whitney was used to evaluate statistical significance. *** = Pvalue < 0.001. Please click here to view a larger version of this figure.
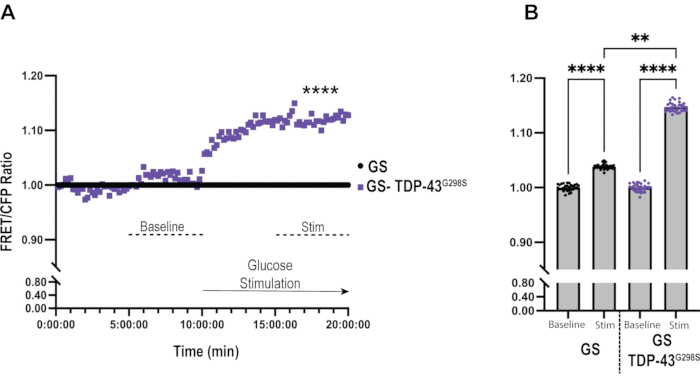
Figure 3: Data analysis of glucose sensor in the context of TDP-43G298S. (A) FRET/CFP ratios of TDP-43G298S are normalized to Glucose Sensor control at each time point. Mann-Whitney was used to evaluate statistical significance. **** = Pvalue < 0.0001. (B) Bar graph representation of glucose uptake in Glucose Sensor control and TDP-43G298S shows a significant increase in glucose uptake in control motor neurons upon stimulation and in the context of TDP-43G298S compared to controls. Kruskall-Walis was used to evaluate statistical significance. ** = Pvalue < 0.01, **** = Pvalue < 0.0001. Please click here to view a larger version of this figure.
