MEVs were purified by differential sedimentation in a density gradient (Figure 1, Figure 2). Under the conditions described, MEVs separated mostly in gradient fraction 3 (F3), which corresponds to 25% iodixanol. This conclusion is based on the detection of protein, membrane lipid, microscopic visualization of intact MEVs, nanoparticle size distribution, and positive reactivity with an antivesicle antiserum (Figure 2, Figure 3). Protein and lipid concentration normalized to colony-forming units (CFUs) showed an approximately eightfold increase of MEV yield in low iron relative to high iron conditions (50 µM FeCl3) (Figure 3). Although the results of one representative experiment is presented, this is a highly reproducible result based on multiple (>10) isolations of MEVs. The pure MEV yield obtained from a 1 L low iron culture by this method was approximately 500 µg of protein.
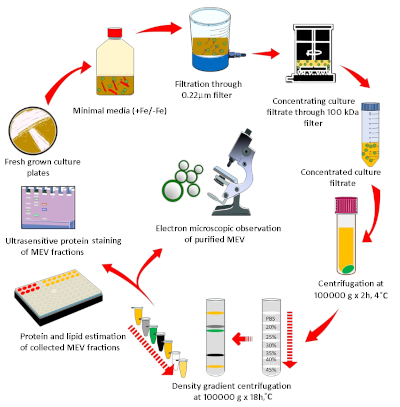
Figure 1: Diagrammatic representation of the methodology used for MEV purification and quantification. Mycobacteria grown in agar plates were used to inoculate iron-depleted minimal medium and grow Mtb for EV isolation. MEVs were purified by a discontinuous density gradient from the cell-free culture filtrate. A combination of membrane lipid and vesicle protein determination, microscopy, and nanoparticle analysis was implemented to characterize purified MEVs. Please click here to view a larger version of this figure.
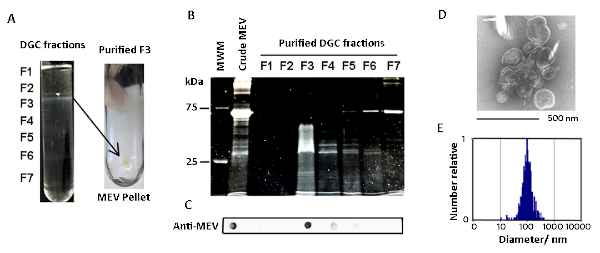
Figure 2: Characterization of purified MEVs. (A) Shown are photographs of an actual density gradient separation of crude MEVs and the pellet of purified MEVs collected by ultracentrifugation of gradient fraction 3 (F3). (B) SDS-gel stained showing the protein profile of the various density gradient fractions. (C) Dot blot analysis showing vesicle-associated proteins concentrated in F3. (D) MEVs present in F3 observed by negative staining. (E) MEV size distribution according to nanoparticle analysis (NTA). Please click here to view a larger version of this figure.
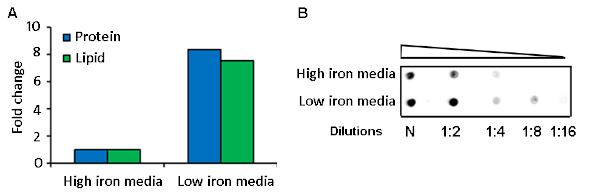
Figure 3: Comparative analysis of MEV yield in low and high iron cultures. A representative result of (A) protein and lipid quantification and (B) dot blot analysis of purified MEVs isolated from iron-limited and iron sufficient Mtb cultures. Please click here to view a larger version of this figure.
