This method allowed characterization of a module of three miRNAs that are consistently downregulated in brain tumors, which are co-expressed specifically during neuronal differentiation (Figure 1) and involved in the tumor survival response after therapy (Figure 2). This is accomplished by regulating a complex oncogenic chromatin repressive pathway. This co-expression pattern suggested a strong synergistic activity among these three miRNAs (Figure 3). Consequently, taking advantage of the small size and simple biogenesis of miRNAs, the second part of this protocol was used to design a transgene (Figure 4) that could simultaneously recapitulate the expression of the three miRNAs in glioblastoma cells, both in vitro and in vivo, with significant interference in tumor biology and promising translational applicability (Figure 5A,B,C)10. Additionally, it was demonstrated that the transgenic cluster is also functional in a breast cancer model (Figure 5D,E).
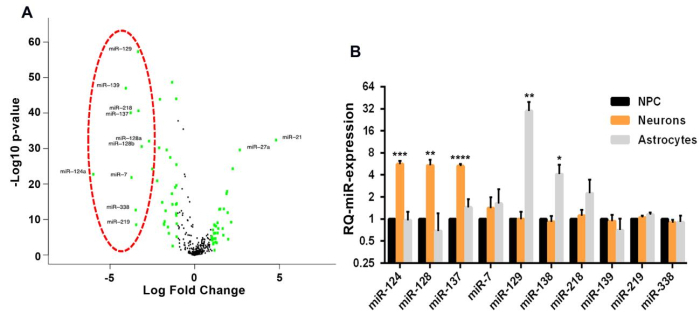
Figure 1: Characterization of a functional miRNA module in glioblastoma. (A) Volcano plot representing the differentially expressed miRNAs in human glioblastoma samples (n = 516) vs. normal brain (n = 10) obtained from TCGA. In green are miRNAs with >4-fold difference. The red circle represents the 10 most significantly downregulated miRNAs in glioblastoma. (B) Relative expression of the 10 miRNAs selected in (A) during induction of different differentiation pathways in neural stem cells, showing clear upregulation of the miR-124, miR-128, and miR-137 modules during induction of neural differentiation. Mean ± SD from three biological replicates (*p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001; Student's t-test, two-tailed). This figure has been modified with permission from reference10. Please click here to view a larger version of this figure.
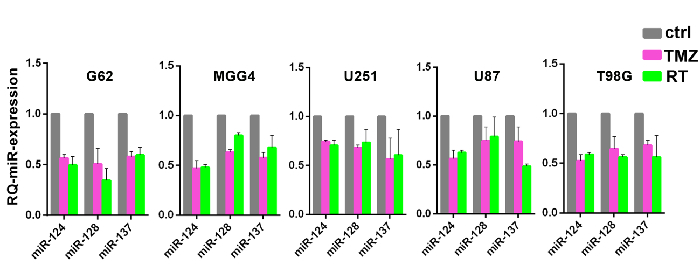
Figure 2: Confirmation of co-expression patterns of miRNA modules during genotoxic stress. Relative expression of the three miRNAs defined in Figure 1 in multiple glioblastoma cells and cell lines (G62-mesenchymal; MGG4-proneural; U251, U87 pro-neural-like, and T98G mesenchymal-like glioblastoma cell lines) after induction of resistance to temozolomide (TMZ, pink bars) or ionizing radiation (RT, green bars). Reported are means with SD from two independent replicates. This figure has been modified with permission from reference10. Please click here to view a larger version of this figure.
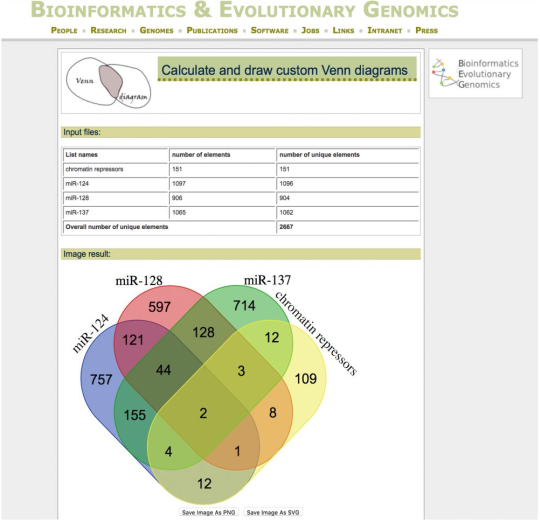
Figure 3: Analysis of functional convergence of the targetomes of co-expressed miRNAs. Venn diagram output from the Bioinformatics and Evolutionary Genomics website, simultaneously crossing the targetome of labelled miRNAs with the mRNAs constituting a GO category of interest (in this case, chromatin repressors). mRNAs uniquely targeted by single miRNAs were chosen for further downstream functional studies. Please click here to view a larger version of this figure.
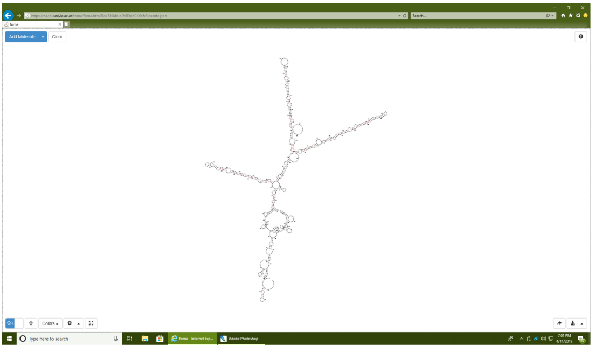
Figure 4: 2D structure of an engineered miRNA sequence encoding the three miRNAs cluster. Graphical output from the RNAweb Fold program. Note the presence of three well defined stem-loop structures which represent the hairpins of each respective miRNA (miR-124, miR-128, miR-137) encoded by the transgene. Please click here to view a larger version of this figure.
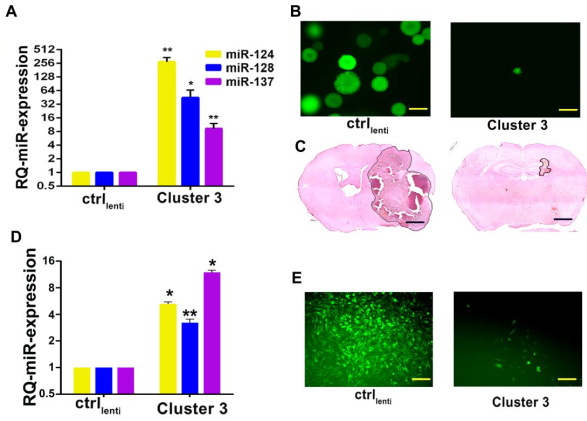
Figure 5: Evidence of transgene processing and its downstream biological effect. (A) Relative quantification of miRNA expression after lentiviral-mediated transduction of G34 glioblastoma cells with the transgenic miRNA cluster (Cluster 3) or negative control (ctrl). Reported are means from three independent experiments ± SD. (B) Fluorescence microscope pictures of G34 glioblastoma spheres expressing negative control transgene vs. Cluster 3 transgene. Scale bar = 100 µm. (C) In vivo growth of intracranial human G34 cell xenografts expressing either control or Cluster 3 transgenes. Scale bar = 1 mm. This figure has been reproduced with permission10. (D) Relative quantification of miRNA expression after lentiviral-mediated transduction of MDA-MB-231 breast cancer cells with the transgenic miRNA cluster (Cluster 3) or negative control (ctrl). (E) Fluorescence microscope images of MDA-MB-231 breast cancer cells expressing negative control transgene vs. Cluster 3 transgene. Scale bar = 100 µm. All experiments were performed in triplicates (*p < 0.05; **p < 0.01; Student's t-test, two-tailed, multiple comparisons). Please click here to view a larger version of this figure.
Supplementary Figure 1: TargetScan workflow. (A) Home page screenshot, showing selection options for miRNA search. (B) Representative search results for miR-137. List of target genes is in the left column. Red box denotes genes with conserved targeting sites (suggesting higher confidence of real targeting). Please click here to view this figure. (Right-click to download.)
Supplementary Figure 2: ToppGene Suite workflow. (A) Home page screenshot showing the search box where the list of genes to be analyzed is inserted. (B) Representative search result for the miR-137 targetome, showing the most statistically significant Gene Ontology (GO) categories. Please click here to view this figure. (Right-click to download.)
