3D Prostate Acini Cultured on Stiff rBM
After 6 days in culture, PECs derived from normal prostate tissue (RWPE-1) (Figure 1A) and BPH tissue (BPH-1) (Figure 1B) form acini on native (PBS treated) rBM that are organized into uniform spheroids of epithelial cells. These acini also have the characteristics of highly organized PECs with apical-to-basal polarity and a visible luminal space13,20.
The acini formed by PECs derived from normal prostate tissue (RWPE-1) (Figure 1A) and BPH tissue (BPH-1) (Figure 1B) on stiffened (AGE-rich) rBM (treated with GLA) have a disrupted architecture (shifting from spheroidal to polygonal in shape and cells protruding/migrating from the acini into the AGE-rich rBM) (Figure 1A). These acini are also characterized by highly disorganized PECs that have lost their apical-to-basal polarity with a small or non-existent luminal space13.
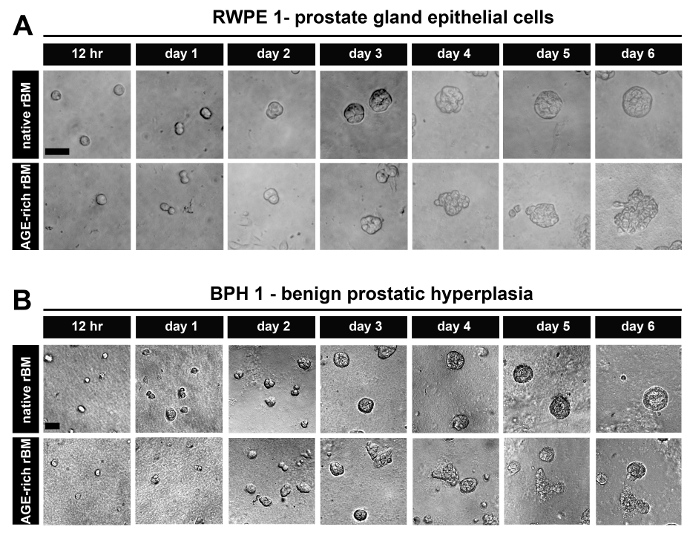
Figure 1: Prostate Epithelial Cells Grown as 3D Glandular Acini on Native and Stiff Reconstituted Basement Membrane (rBM). (A) Brightfield images of RWPE-1 cells grown for 12 hr up to 6 days on rBM gels treated with PBS (native) or 50 mM glycolaldehyde for 14 hr (AGE-rich; stiff); Scale bar = 50 µm. (B) BPH-1 cells, grown as described in panel A; Scale bars = 50 µm; data is representative of 3 independent experiments. Please click here to view a larger version of this figure.
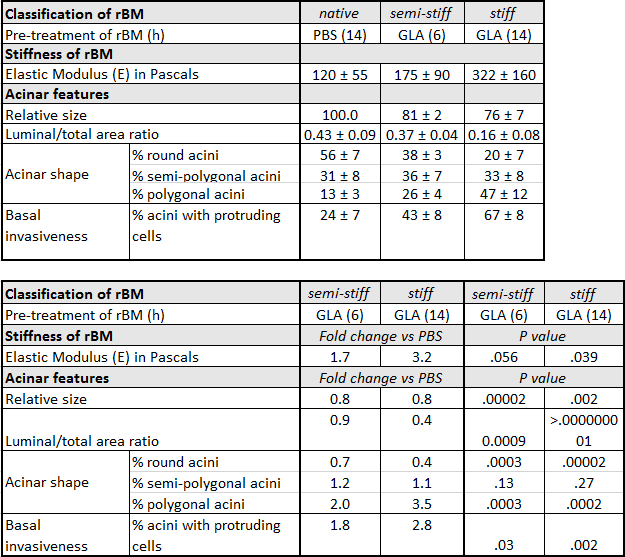
Table 1: Characteristics of Prostate Epithelial RWPE-1 acini Grown on Native, Semi-Stiff and Stiff Reconstituted Basement Membrane (rBM). RWPE-1 acini were grown on rBM pre-treated with PBS for 14 hr (native), glycolaldehyde (GLA) for 6 hr (semi-stiff) or GLA for 14 hr (stiff). For acinar shape, the percentage (%) ± standard deviation (SD) of round, semi-polygonal and polygonal acini were calculated from 5 independent experiments (50 acini quantified per condition). Relative acinar size was calculated (native rBM = 100 %) from 3 independent experiments. For invasiveness, % ± SD acini with one or more protruding cells were calculated from 3 independent experiments. Fold change is calculated by dividing the average value obtained under semi-stiff or stiff conditions by the corresponding value for native conditions. P values calculated using Student's t-test (α = 0.05).
AGE dependent increased rBM stiffness promotes PC3 prostate tumor cell migration
PC3 cells grown on native rBM migrate by maintaining continuous cell-cell contact, whereas PC3 cells grown on AGE-rich (stiff) rBM move independently from each other (Figure 2A). After 72 hr in culture PC3 cells form foci (spheroids) on native (PBS treated) rBM, whereas PC3 cells on stiff (AGE-rich) rBM do not from spheroids and migrate independently (Figure 2B). PC3 cells on stiff (AGE-rich) rBM are more elongated than PC3 cells grown on native rBM (Figure 2C). PC3 cells on stiff rBM migrate faster than PC3 cells grown on native rBM (Figure 2D). PC3 cells on stiff rBM display a decrease in persistence compared to PC3 cells grown on native rBM (Figure 2E).
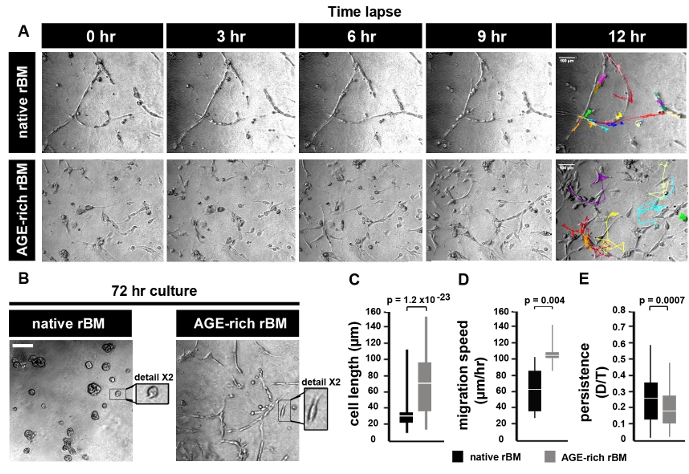
Figure 2: Prostate Tumor Cell Migration on Native and Stiff Reconstituted Basement Membrane (rBM). (A) Brightfield images of PC3 cells grown on rBM gels treated with PBS (native) or 50 mM glycolaldehyde for 14 hr (AGE-rich, stiff). Cells were imaged using a brightfield microscope (10X objective) and an acquisition rate of 1 image per hr for 12 hr followed by cell tracking to generate trajectories. Images shown correspond to the time points after 0, 3, 6, 9 and 12 hr. Trajectories of single cells are shown for the 12 hr time point. Scale bar = 100 µm. (B) PC3 cells cultured on native or stiff rBM for 72 hr, and imaged as described in panel (A). Scale bar = 100 µm. Detail shows selected area at 2X magnification. (C) Mean ± S.D. cell length (µm); significant difference between native rBM and stiff rBM (p = 1.2 x 10-23). (D) Mean ± S.D. velocity (µm/hr) calculated from cell trajectories; significant difference between native rBM and stiff rBM (p = 0.004). (E) Mean ± S.D. persistence of cell movement (ratio D/T, where D = distance from start to end of cell trajectory, T = total length of cell trajectory); significant difference between native rBM and stiff rBM (p = 0.0007). For panels C-E >10 cells were analyzed, data is representative of 3 independent experiments. Please click here to view a larger version of this figure.
A protocol for the generation of 3D glandular acini from MECs in pure rBM gels6 was modified in a previous study by the addition of 4 mg/ml type I collagen to the rBM matrix. The addition of collagen resulted in the elastic modulus of the rBM gel increasing from 175 ± 37 to 1589 ± 380 Pascals. This 9.1-fold increase in stiffness modulated the growth, survival, migration and differentiation of MECs21. The protocol was modified again by including a treatment step with D-(-)-ribose to promote non-enzymatic crosslinking of the type I collagen that had been added to the rBM gel. The resultant 15-fold increase in stiffness was found to cooperate with oncogenic transformation of MECs to promote their invasive behavior22. The experimental approach of adding type I collagen to rBM gels facilitates the direct interaction of MECs with collagen fibres, which only occurs in human tissue after the physical barrier between the stroma and epithelium provided by the BM undergoes proteolytic degradation. By generating 3D glandular acini from PECs in pure rBM gels pre-treated with GLA, the current protocol opens the way to study how BM stiffness per se can trigger their invasive behaviour (Figure 3). The levels of BM stiffness induced in this protocol have physiological relevance. Incubation with 50 mM GLA for 6 hr and 14 hr respectively increased the elastic moduli of the pure rBM gel to 175 ± 90 and 322 ± 160 compared to 122 ± 55 Pascals in rBM gels treated with PBS (Table 1). This 1.7 to 3.2-fold increase in rBM stiffness recapitulates the 2.5- to 3.4-fold increase in stiffness observed in malignant compared to normal prostate or BPH tissue23-26. As outlined in a recent publication13 the morphological changes induced by the accumulation of AGE and rBM stiffness in PEC acini can be quantified for a statistically significant shift from a rounded to polygonal shape, decreased luminal/total acinar area, and protruding cells migrating from the acina into the AGE-rich rBM (Figure 3). Immunoblotting can also be used to assess markers of EMT (e.g. loss of E-cadherin13) and the contractile behavior (e.g. phosphorylated myosin light chain-2, pMLC213) in PECs grown in normal versus stiff rBM (Figure 3). Further evaluation using immunofluorescent staining and confocal microscopy can be applied to visualize the BM (e.g. laminin, collagen IV and AGE accumulation13), cellular apical-to-basal polarity (e.g. apical localization of EEA1: early endosomal antigen 1; and GM130: 130 kDa cis-Golgi marker13) and cellular patterns of adhesion molecules (e.g. E-cadherin localization to cell-cell junctions13) (Figure 3).
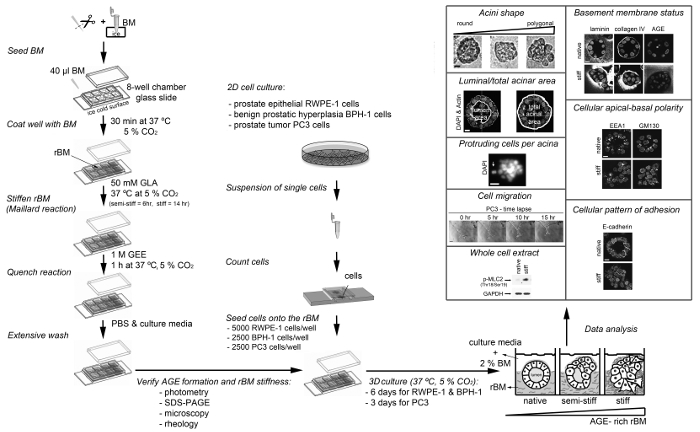
Figure 3: Overview of the Different Protocols Presented Here. The diagram depicts how to prepare and stiffen the reconstituted basement membrane (rBM) with glycolaldehyde (Maillard reaction), how to seed cells on to the stiff rBM, how to analyze the stiff rBM (extent of Maillard reaction) and procedures that can be used to analyze the cellular and molecular changes induced by AGE-rich rBM. AGE, advanced glycation endproducts; BM, basement membrane; DAPI, 4',6-diamidino-2-phenylindole; EEA1, early endosomal antigen 1; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; GLA, glycolaldehyde; GEE, glycine ethyl ester; GM130, 130 kDa cis-Golgi marker; p-MLC2 (Thr18/Ser19), myosin light chain-2 phosphorylated at sites threonine 18 and serine 19; rBM, reconstituted basement membrane; SDS-PAGE, sodium dodecyl sulfate polyacrylamide gel electrophoresis. For RWPE1 acini Scale bar = 10 µm; for PC3 tumor cell spheroids Scale bar = 100 µm. This figure has been modified from reference13. Please click here to view a larger version of this figure.
Troubleshooting steps will be necessary if D-(-)-ribose is chosen as the crosslinking agent for rBM. During protocol development it was found that treatment with 1 M D-(-)-ribose for 72 hr, as previously described for rBM/collagen gels22, resulted in the dehydration and shrinkage of rBM gels. The evaluation of lower concentrations of D-(-)-ribose and shorter treatment times may help to overcome this limitation.
A potential limitation in future applications of the protocol could be encountered where higher levels of rBM stiffness are desired. If longer incubation times and higher concentrations of GLA are used to induce higher levels of rBM gel stiffness it will be necessary to assess whether these treatment conditions have an impact on cell survival and proliferation, as previously described13. It should also be noted that incubation of RWPE-1 cells with serum induces a phenotypic EMT-like transition and exposure to serum or serum-containing materials should be avoided. For example, if experiments involve the transfection of short interfering RNA (siRNA) oligonucleotides, the procedure should be optimized using RWPE-1 cells grown in KSFM, without switching the cells to low serum transfection media. This drawback could compromise the level of gene silencing achieved when using transient siRNA approaches in the model. For some protein targets it would be advised to employ inducible shRNA vectors for tunable gene silencing and the desired decrease in protein levels. Adaptations that incorporate enzymatic crosslinking by stromal cell or tumor cell associated lysyl oxidase (LOX)17 could also be incorporated into future models.
This protocol will facilitate the future study of pro-invasive mechanisms triggered by AGE-dependent BM stiffness in PECs (RWPE-1, BPH-1) and evaluation of anti-metastatic targets in invasive PTCs (PC3). Given that BPH is considered to be a metabolic disorder27, this protocol also paves the way towards our improved understanding of the link between metabolic disorders and increased prostate cancer risk. Given that BM stiffness induced by its exposure to AGEs may be a trigger for invasiveness in other cancer types, it will be of interest to use the protocol to set up similar models that incorporate normal epithelial cells and tumor cells from other organs (e.g. breast, colon, ovaries, pancreas).
Critical steps within the protocol, together with their timings, are summarized in Figure 4. During the initial step it is essential to maintain the stock solution of rBM at 4 °C while it thaws to prevent its polymerization. Pipette tips should not be placed into the rM stock solution until they have been chilled to 4 °C. For the next step it is also important to ensure the chamber slides have equilibrated to 4 °C before they are coated with the rBM solution. As soon as the temperature of the rBM solution is increased above 4 °C it will undergo irreversible polymerization to form a gel. It essential that the rBM is not disturbed during the polymerization stage to ensure that it forms an even surface suitable for cell culture and microscopic analysis. The duration of incubation with GLA with or without inhibitors of the Maillard reaction (sodium cyanoborohydride and amingoguanidine) will determine how stiff the rBM gel becomes. It is recommended to use a 6 hr incubation with GLA if semi-stiff conditions are required, and 14 hr incubation if stiff conditions are required (Table 1). Alternate incubation times or concentrations of GLA can be used if different levels of stiffness are desired. In this case rheological analysis of the rBM gels need to be incorporated as an essential step. Following the step of quenching the Maillard reaction by incubation with GEE and the subsequent washing steps with PBS, the rBM gels can be used immediately or stored at 4 °C for up to 48 hr prior to their use for cell culture. Once cell cultures are set up it is important to change the culture medium (including any treatments) every two days. It is recommended to maintain the 3D cell cultures for 3-12 days according to the parameters under investigation. For 3D PEC acini it is recommended to analyse the cultures after 6 days, and for 3D PTC spheroids analysis is recommended after 3 days of culture in the first instance.
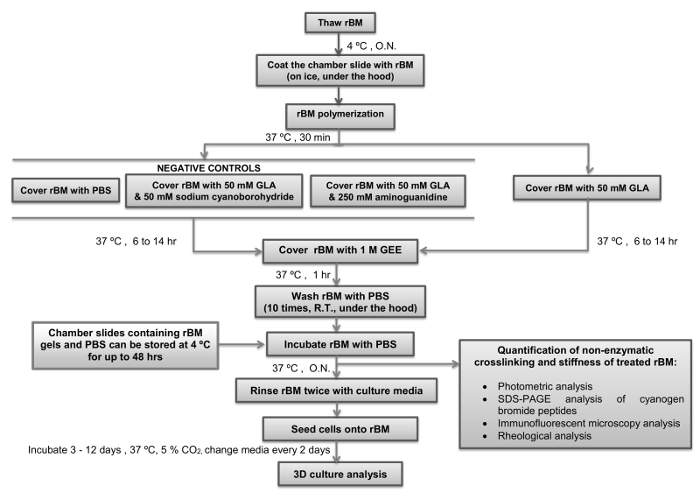
Figure 4: Simple Overview of the Protocol with Critical Steps and Timings Indicated. The flow diagram depicts how to prepare and stiffen the reconstituted basement membrane (rBM) with glycolaldehyde (Maillard reaction) with critical steps and timings indicated. Points where the protocol can be stopped, and rBM gels stored, are also indicated. rBM, reconstituted basement membrane; GLA, glycolaldehyde; GEE, glycine ethyl ester; O.N., overnight; PBS, phosphate buffered saline; R.T., room temperature. Please click here to view a larger version of this figure.
