Enrichment of Sca1high ASCs from Different Fat Pads.
The vascular stromal cells isolated from SQ fat display fibroblast-like, stretched cell shape regardless of Sca1 expression level (Figure 1A). On the other hand, VIS (eWAT-derived) Sca1high and Sca1low cells demonstrate distinct difference in their cell shape. Like SQ (iWAT-derived) Sca1high cells, VIS (eWAT-derived) Sca1high cells display stretched, fibroblast-like cell shape, whereas VIS Sca1low cells demonstrate epithelioid shape. Sca1high cells isolated from Sca1-GFP mice are easily identified as GFP-positive cells in tissue culture (Figure 1B). When these cells were assessed with flow cytometry, most of the GFP positive cells were confirmed to express Sca1 proteins on the cell surface as detected with Anti-Sca1 antibody (Figure 1C). Sca1high cells derived from inguinal fat pad maintain increased capacity of adipocyte differentiation whereas eWAT-derived Sca1high cells are more difficult to differentiate into adipocyte with conventional adipogenic mix10.
Fat Depot-dependent Gene Expression of Sca1highASCs (Figure 2).
Genome-wide transcriptome analyses with RNA sequencing demonstrated the enrichment of genes related to extracellular matrix proteins and modifiers (GO:0031012, GO:0005578) in those Sca1high ASCs10. Coupled with real-time PCR analyses, we were able to demonstrate the differential expression of the collagenolytic MMPs (MMP2, MMP8, MMP13, MMP14) between iWAT- and eWAT-derived Sca1high ASCs. When fluorescein-labeled type I collagen gels were used to assess pericellular degradation activity, we observed the markedly increased collagen remodeling activity mediated by VIS Sca1high ASCs10.
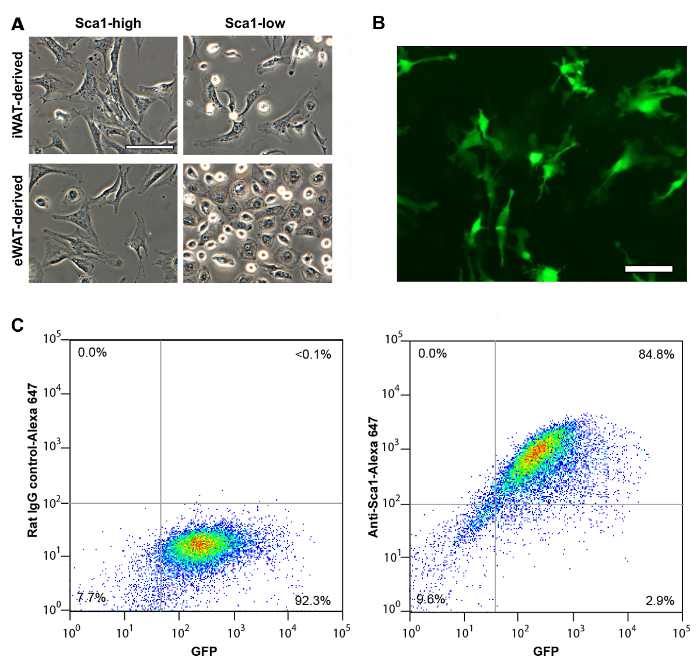
Figure 1: Immunomagnetic Separation of Murine Sca1high ASCs from Different Fat Depots. (A) Sca1high and Sca1low ASCs isolated from SQ (iWAT) and VIS (eWAT). Scale = 100 µm. (B) Sca1-GFP cells isolated from iWAT of Sca-GFP mice. Scale = 100 µm. (C) Cell surface expression of Sca1 in Sca1-GFP-positive cells assessed with flow cytometry. (left) control rat IgG (right) anti-Sca1 antibody. X-axis, GFP intensity. Y-axis, Alexa-Fluor 647. Panel A shown previously in Tokunaga, M. et al., (2014). Please click here to view a larger version of this figure.
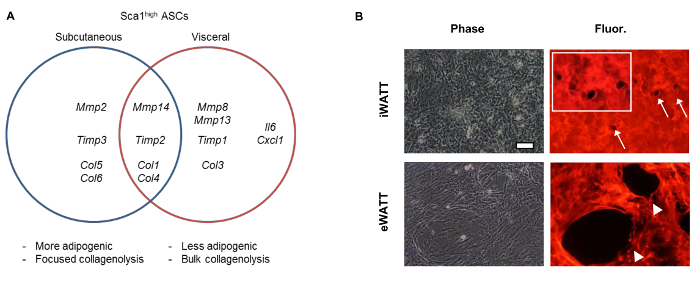
Figure 2: Fat Depot-Dependent Expression of Collagenolytic MMPs, TIMPs, and Collagens. (A) Differential gene expression of ECMs and ECM modifiers in Sca1high ASCs isolated from different fat depots. (B) Increased collagen degradation activity of VIS (eWAT-derived) Sca1high ASCs. Degradation of collagen is shown as the disappearance of fluorescent signals (arrows and arrowheads). Inset is an enlarged image of focused collagen degradation mediated by individual cell of SQ ASCs. Cells were cultured for 72 hr. Data shown previously in Tokunaga, M. et al., (2014). Please click here to view a larger version of this figure.
