1. Growing Seedlings and Preparing Bacteria
- Seedlings are germinated in MetroMix 500 soil mixture (BWI Companies) in a growth chamber (160 μmol photons m-2s-1 with a photoperiod of 8 hour light at 22 °C and 16 hour dark at 20 °C for 1 week. Relative humidity was set to 70%. Seedlings were watered with tap water as needed to keep the soil from becoming dry.
- Two seedlings of similar size are then transplanted to 4″ diameter pots containing the MetroMix 500 soil and grown for an additional 3 weeks in the same conditions as 1.1.
- Pseudomonas syringae (Pst) is initially T-streaked onto Kings B Medium (KBM) plates (per liter: 20 g Proteose Peptone, 0.1975 g MgSO4, 1% Glycerol, 1.5 g K2HPO4, 18 g Agar) and grown for two days at 28 °C. A single colony is collected, mixed into 300μl liquid Kings B Medium (20 g Proteose peptone, 0.1975 g MgSO4, 10 mL Glycerol, 10 mL of 100x K2HPO4 stock (1.5 g of K2HPO4 in 10 mL of H2O ), and spread onto a Kings B Medium plate. This is cultured over night at 28 °C.
- The bacteria inoculum is prepared by scraping the cultured bacteria from the KBM plate into 20 mL H2O. An OD600 of 0.03 (~106 cfu/ml) is prepared in 1 L of inoculation buffer (10 mM MgCl2 + 400 μL Silwett-70). Inoculation buffer minus bacteria was prepared for the control solution.
2. Inoculation of Tomato with Pst and H2O2 Histochemical Stain
- Four week old plants were inoculated by dipping into the above inoculation solution for 30 seconds. Clear plastic bags were put over the plants immediately after inoculation.
- 3′-3′ diaminobenzidine (DAB) stain11 was prepared (1 mg/ml DAB in H2O, pH 3.8 (HCl)) three days before use to achieve complete solubilization. The solution was rocked to mix at room temperature.
- Twenty four hours after treatment, the fully expanded leaf was removed, placed in DAB stain epidermal side up, and vacuum infiltrated for 15 min. The leaves were then put in the dark at room temperature over night for staining.
- Leaves were boiled in 95% ethanol for 10 min, and then kept in 75% ethanol.
3. Protein Extraction
- After harvesting, tomato leaves were ground to fine powder with liquid nitrogen using mortar and pestle.
- For 0.5 g of tissue, add 1.25 mL Tris pH 8.8 buffered phenol and 1.25 mL / 0.5 g of extraction buffer (0.1 M Tris-HCl pH 8.8, 10 mM EDTA, 0.9 M sucrose) then continue grinding for a few more min in a fume hood.
- Transfer the extract to Oakridge centrifuge tube (Thermo Fisher Scientific Nalgene Company) and agitate for 2 hrs at room temperature.
- Centrifuge at 5,000x g and 15 °C for 10 min. Transfer the top phenol phase to a new clean tube.
- Back-extract the aqueous phase with equal volume extracted phenol buffer; agitate on a shaker for 30 min. Centrifuge and transfer the extract to a new Falcon tube.
- Repeat the above step one more time and transfer the extract to new Oakridge tube.
- Precipitate phenol extracted proteins by adding 5 volumes of 0.1 M ammonium acetate in 100% methanol (stored at -20 °C). Vortex and incubate at -20 °C for overnight.
- Collect the protein pellet by centrifugation at 20,000x g, 4 °C for 20 min .
- Wash the pellet 2 times with the cold 0.1M ammonium acetate in methanol and 2 times with 80% cold acetone. Add 1.5 mL cold 70% ethanol to pellet and suspend. Transfer to a 2 mL microcentrifuge tube (USA Scientific)and centrifuge at 14,000 rpm, 4 °C for 20 min. Remove ethanol.
- Dry the pellet briefly in a SpeedVac concentrator and dissolve in a protein extraction buffer, e.g. ReadyPrep Protein Extraction Kit Reagent 3 (8 M Urea, 4% CHAPS, 40 mM Tris-base, 2 M Thiourea) (Bio-Rad). Centrifuge at 14,000 rpm and 20 °C for 30 min to form a pellet. Collect the supernatant.
- Measure the protein concentration using a CB-X protein assay according to manufacturer’s manual (Geno Technology).
4. Sample Preparation and Peptide Labeling with cysTMTs
- Prepare protein sample at a concentration of 2-5 μg/μL. Here we use 100 μg of protein for each mass tag, 20 μl-50 μL samples. For this experiment all 6 tags were used.
- Add equal volume of alkylation buffer (100 mM Tris-HCl pH7.5, 200 mM iodoacetamide) to the protein sample to block the free thiol group. Buffer should be prepared fresh. Alkylation is performed at 37 °C for 1 hour.
- Precipitate the protein by adding 1 mL of cold 80% acetone at -20°C for overnight. Pellet the protein by centrifuging at 14,000 rpm, 4 °C for 20 min. Wash the pellet 3 times with 80% Acetone vortexing each time. Remove the supernatant and allow to air dry while on ice.
- Resuspend the pellet in 50 μL lysis buffer (6 M Urea, 50 mM Tris base, 1 mM EDTA). 0.5 μL of 100 mM Tris(2-Carboxyethyl) Phosphine (TCEP) is then added to reduce the disulfide bonds by incubating for one hour at room temperature.
- Prepare tags by adding 20 μL of acetonitrile to the reagent tubes to solubilize the tags. Vortex and spin down to the bottom.
- Wash out extra TCEP using a Microcon 3KD centrifugal filter device (Millipore). Add 50 μL sample to Microcon 3KD column, and 50 μL lysis buffer to the column. Centrifuge for 15 min at 10,000x g and 4 °C. Repeat this step for a total of three times. Remove column and invert into a clean tube. Centrifuge for 5 min at 1,000x g and 4 °C.
- Check the pH. If needed, adjust the pH to 7.0-8.0 with 1 M HCl.
- Add 5 μL of the cysTMT reagent to each sample (The cysTMT reagent kit provides 20 μL tags for up to 500 μg protein. This method uses 100 μg protein, therefore we use one-fifth of the tag.) . Allow the reaction to proceed for 2 hr at room temperature.
- Samples were combined with Laemmli Sample Buffer (Bio-Rad) in a 1:1 ratio.
5. Removal of Non-reacted Tag Sample Fractionation
- Samples were boiled for 5 min and separated on a pre-cast 12% polyacrylamide gel (Bio-Rad). The gel was rinsed three times in 5 minute intervals with filtered H2O and stained with Coomassie Blue (Bio-Rad) for 1 hour. The gel was de-stained overnight with H2O.
- Twelve fractions were collected from each gel lane. Fractions were determined by protein band concentrations. Fractions were chopped into 1mm pieces and collected into a 1.5 mL tube.
- Gel pieces were de-stained using 50% acetonitrile and 0.1 M ammonium bicarbonate in H2O. This step must be repeated until blue color is removed from gel pieces (~3-4 times, 15 min each time).
- De-stained gel pieces were dried using 100% acetonitrile (cover gel pieces) for 15 min, supernatant removed, and the pieces dried using a SpeedVac.
- Dissolve trypsin (Promega) at 1:10 (trypsin: protein) ratio in the same volume (as pooled labeled sample) of 25 mM ammonium bicarbonate. For example, 600 μg protein sample with a volume of 500 μL needs 60 μg trypsin dissolved in 500 μL of 25 mM ammonium bicarbonate.
- Add the trypsin solution to the gel pieces and set on ice to rehydrate. If pieces are not covered add 50 mM ammonium bicarbonate buffer. After rehydration, incubate at 37 °C for 12-16 hr.
- Remove the liquid protein extract from the incubated samples.
- Stop the reaction and remove any remaining protein digest by covering gel pieces with 5% formic acid, 50% acetonitrile.. Shake gel pieces for 20 min at room temperature. Transfer the supernatant to the extracted protein sample tubes from 5.7. Repeat the extraction procedure twice. Dry the extracted peptides using the SpeedVac.
6. Sample Enrichment of cysTMT Labeled-peptides
- Add a gradient of anti-TMT resin to 0.5 mL tubes for a 50% slurry. (Gradient was determined from band concentration in fractionation gel. If a fraction collection consists of a dark band, more resin is needed. Fractions with weaker bands require less resin as there is less protein.)
- The resin is then washed three times with one column volume of 1x Tris-buffered saline (TBS) (25 mM Tris, 0.15 M NaCl, pH 7.2) (Thermo Scientific Pierce Protein Research Products).
- Add 200 μL 1xTBS to each sample. Samples are then added to anti-TMT resin (Thermo Scientific Pierce Protein Research Products) and agitated at room temperature for 2 hr followed by rocking overnight at 4 °C.
- Add sample to column (Thermo Scientific Pierce Protein Research Products).
- Wash each column three times with 1x 200 μL TBS. This is then followed with washing three times with 0.05% CHAPs (dissolved in 1x TBS).
- The column is then washed three times with 4 M urea in 1x TBS. Two hundred microliters of H2O is then used to wash the column three times.
- Each sample is eluted three times with 200 μL 50% elution buffer (50% acetonitrile, 0.4% TFA).
- Samples are then dried in a vacuum concentrator.
7. Mass Spectrometry Analysis
- Resuspend the samples in 12 μL of 3% acetonitrile with 0.1% formic acid and inject 5 μL directly onto an Eksigent NanoLC-1D high pressure liquid chromatography column (AB SCIEX, USA).
- Peptides will be separated on a Proteopep II C18 column 75 μm ID x 20 cm (New Objective, USA) using a 4-60% gradient (A: 3% acetonitrile, 0.1% formic acid; B: 97% acetonitrile, 0.1% formic acid) at 3 μL/min over 60 min.
- A Thermo Scientific LTQ Orbitrap XL mass spectrometer was used to detect peptides using a top 2 x 3 experiment consisting of single stage MS followed by acquisition of 3 MS/MS spectra with higher-energy C-trap dissociation (HCD) fragmentation followed by 3 MS/MS with collision induced dissociation (CID) for protein identification. Parameters in this mode were: Isolation width: 3.0 m/z; Collision energy: 50% (10% with two steps). Only doubly- and triply-charged peptides were selected for fragmentation. Dynamic exclusion parameters were set to: Repeat count = 1; Repeat Duration = 60; Exclusion list size = 500; Exclusion duration = 28. Target values are as follows: MS = 5 e5; MS/MS (HCD) = 1 e5. Ion transfer times were set to 500 for FTMS and 300 for MS/MS (HCD). Two microscans were required for HCD spectra.
8. Database Searching and Quantitation
- The acquired CID and HCD data were analyzed using Thermo Scientific Proteome Discoverer software 1.2 (Thermo Scientific Pierce Protein Research Products) through a branched workflow which implemented a Reporter Ion Quantizer (20 ppm mass tolerance of fragment ions) to quantify the ratio. A separate segment processed the MS2 spectra through the Spectrum Selector, Spectrum Normalizer, and Spectrum Grouper nodes.
- Data were then searched against the SEQUEST search engine. A 20 ppm mass tolerance was used. Static modifications were set to the cysTMT reagent (304.18 Da) and dynamic modifications included phosphorylation and methionine oxidation (15.99 Da). A custom database for Solanum lycopersicum was composed using RNA data (with about 350,000 entries from Harvard University)13 was used in both cases.
9. Representative Results
A representative image of a control tomato plant leaf and a Pseudomonas inoculated leaf is shown in Figure 1. A difference between control treated and Pseudomonas treated leaves is observed. After the leaves are removed and stained using DAB, the de-staining process allows for the histochemical stain to show signs of ROS in the leaf tissue (Figure 2). Figure 2A is representative of a control leaf with no staining. Figure 2B is representative of a leaf treated with Pseudomonas and positive staining for H2O2 production. An example of Proteome Discover data output of a differentially redox regulated protein is shown in Figure 3. This protein is a known redox regulated protein ferredoxin-114 and has been shown to play a role in defense against Pseudomonas syringae pv tomato15. Peak intensity between control and inoculated samples is used to obtain relative quantification, which showed significant changes in ferredoxin-1 redox regulation (p< 0.05). High intensity peaks suggest that this protein is oxidized in response to the pathogen treatment. Figure 4 is an example of Proteome Discover data output of a protein that has similar redox regulation of a protein between a control and inoculated sample. Peaks of similar intensity suggest the presence of disulfide bonds not regulated by a change in treatment. The method will revolutionize how scientists detect redox responsive cysteines and disulfides10.

Figure 1. A representative image of inoculated tomato leaves with control solution (A) and Pseudomonas (B).

Figure 2. A representative image of DAB staining of inoculated tomato leaves with control solution (A) and Pseudomonas (B). Leaves were stained using 3′-3′ diaminobenzidine. Chlorophyll was removed from leaves by boiling in 95% ethanol. Dark staining indicates the presence of H2O2. Only leaves inoculated with bacteria culture showed dark staining.
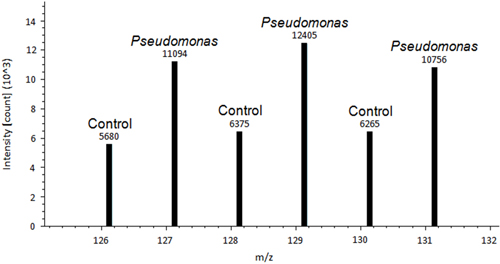
Figure 3. An example of Proteome Discover data output of ferredoxin-1, differentially redox regulated protein14. Peak intensity above each peak is used for absolute quantification. Peak intensity between control and inoculated samples is used to perform relative quantification.
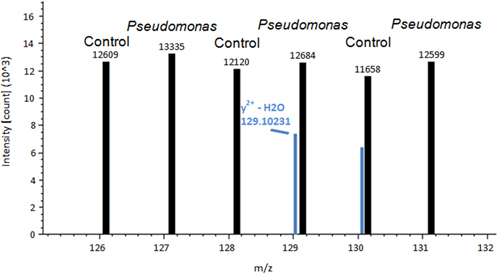
Figure 4. An example of Proteome Discover data output of a protein that has similar peak intensity between a control and inoculated sample. Peaks of similar intensity suggest the presence of disulfide bonds not regulated by a change in treatment.
