1. Vaginal inoculation with Candida albicans
- Three days prior to inoculation, while restraining the animal to expose the abdomen, inject 100 μl of sesame oil containing 0.1-0.5 mg of β-estradiol subcutaneously in the lower abdomen. Advance the needle about 5 to 10 mm lateral to the skin to minimize leakage from the injection site.
The subcutaneous administration of estrogen in the lower abdomen is optimal in this model due to the close proximity to the genital tract. Effective doses may vary by mouse strains, ages or estrogen derivatives. In previous studies using CBA-J (H-2κ), C3H/HeN (H-2κ), C57BL/6 (H-2b), Balb/c (H-2d), DBA/2 (H-2d), SJL (H-2s) mice at 6-8 weeks of age, 0.1 mg/100 μl was found effective evidenced by thickening of the vaginal wall, reduced vaginal mucus and increased epithelial cell sloughing. Mice above treated with estrogen at this concentration exhibit consistent vaginal colonization with Candida. For inoculation in mice of other strains and ages, a pilot study is recommended to ensure the effectiveness of estrogen under the modified conditions and increase the estrogen dose if necessary.
The estrogen solution should be prepared fresh each time on the day of injection. To ensure complete solubility of estrogen in sesame oil, thoroughly mix the solution using a vortex mixer and heat intermittently at 37°C. Repeat the injection weekly throughout the study period. - To prepare the inoculum, add a loopful of C. albicans blastoconidia from a recent subculture preparation on Sabouraud-dextrose agar (SDA) into 10 ml of Phytone-peptone broth supplemented with 0.1% glucose. Incubate the broth culture to stationary phase for 18 h at 25°C in a shaking water bath.
- Following incubation, collect the broth culture into a 15 ml conical tube and centrifuge at 800 x g for 5 min. Wash the pellet twice with sterile PBS.
- Enumerate viable blastoconidia on a hemocytometer by trypan blue dye exclusion. Adjust the cell concentration to 2.5 x 106/ml (or to a desired inoculum concentration) in sterile PBS.
- To stabilize the mouse, hold the base of the tail with two fingers and lift the hip upward so that the vaginal opening faces toward you (Figure 1A). It is ideal if the mouse is placed on a flat grated surface (e.g. cage top) so that the mouse can provide resistance against the tail restraint.
- Pipette 20 μl (or a desired volume not to exceed 20 μl) of the inoculum suspension by inserting the pipette tip about 5 mm deep into the vaginal lumen (Figure 1B). Complete this step as quickly and gently as possible to minimize distress in the mouse.
2. Vaginal lavages
- Following euthanasia (or anesthesia), hold the mouse downward by the base of the tail with two fingers so that the vaginal opening becomes exposed.
- Lavage the vaginal lumen by introducing 100 μl of sterile PBS with repeated aspiration and agitation with a pipette tip. The pipette tip may become clogged with cells. If this occurs, dispense the obstructing cells and continue lavaging with remaining PBS in the vagina. Collect the lavage fluid into a microcentrifuge tube.
- Alternatively, vaginal lavages can be performed on anesthetized mice with isoflurane inhalant anesthesia. For this, expose the mice to vaporized isoflurane until they are fully sedated (~30 sec.). Hold the mice downward by the base of the tail and gently lavage the vaginal lumen using 50 μl of sterile PBS. Make sure to avoid harsh agitation with a pipette tip to minimize trauma to the vagina during this procedure. Sedated mice should recover from anesthesia within 30 sec of exposure to ambient air.
Isoflurane can be vaporized using an isoflurane vaporizer and O2 (preferred) or a standard drop system closed anesthetic chamber without the vaporizer system (requires close monitoring of the animal while sedated to avoid respiratory distress).
Vaginal lavages on anesthetized mice should be the method of choice for longitudinal lavage samples on the same mice. Consecutive lavaging does not influence assessment of fungal burden over time41. - For wet-mount preparations, transfer 10 μl of the lavage fluid onto a glass slide and observe at 400-1000x magnification by light microscopy. Additionally, cellular fractions of the lavage fluid can be stained to examine cell and nuclear morphologies. For smear preparations, transfer 10 μl of the lavage fluid onto a glass slide and gently spread using the outer wall of a pipette tip. Preserve the smear samples with CytoPrep fixative and stain by the standard Papanicolaou technique (Pap smear). Observe at 400 × magnification by light microscopy.
3. Quantification of vaginal fungal burden
- In a 96 well round-bottom plate, transfer the lavage fluid into one well of the top row and 180 μl of sterile PBS into the following 5 wells of that column (down the plate).
- Make 1:10 serial dilutions of vaginal lavage fluid by transferring the 20 μl of the fluid to the next well in the column. Mix thoroughly by repeated aspiration before each transfer. Serial dilutions of up to 12 lavage samples (one full horizontal row) can be performed simultaneously using a 12-channel pipette.
- Starting with the lowest dilution, transfer 10 μl of the sample onto Sabouraud-dextrose agar (SDA). Plating of up to 36 samples can be performed on 1 plate using SDA prepared in square petri dishes with grid and an adjustable spacing multichannel pipette.
- Enumerate colony forming units (CFUs) after incubation at 34°C for 48 h.
4. Vaginal tissue extraction
- Following the vaginal lavage procedure, lay the euthanized mouse on its back and saturate the groin area with 70% ethanol. Using a pair of forceps, lift the urinary orifice upward so that the vaginal opening becomes exposed.
- Insert a pair of curved forceps into the vaginal lumen and locate the cervix. While maintaining a firm grip with the forceps, extract the cervix through the vaginal cavity (Figure 2A-C).
- Excise the vagina at the base of the vaginal opening and then remove the cervix from the vagina with surgical scissors. Keep in mind that the vaginal tissue is involuted (inner epithelium-side of the vagina is exposed outward). The tissue can be either laterally inverted to maintain the original orientation of the vagina or opened into a sheet by making a lateral incision.
Excised vaginal tissues can be used for 1) lymphocyte extraction following collagenase digestion (~1 ×104/mouse)40, 2) epithelial cell isolation following dispase digestion (~5 ×104/mouse)28, 3) frozen or paraffin-embedded preparations for histological analyses25.
5. Lumbar lymph node excision
- Following the vaginal lavage procedure, lay the euthanized mouse on its back and saturate the abdomen with 70% ethanol. Make a lateral incision starting from the lower abdomen to the chest and expose the internal organs. Using a pair of forceps in both hands, move the intestines upward so that the central blood vessels become visible.
- Locate the inferior vena cava and abdominal aorta. Normally, a pair of lumbar lymph nodes can be identified adjacent to the abdominal aorta, located about halfway between the origin of the renal and common iliac arteries39. These lymph nodes can be visually distinguished from fat tissue by the elastic texture and are lighter and more opaque in color compared to fat tissue (Figure 3). These lymph nodes are noticeably larger in infected animals compared to uninoculated animals.
- Excise the lymph nodes by placing microforceps under the node and then pull up gently to separate from surrounding tissue.
6. Isolation of lymphoid cells in single-cell suspensions
- Transfer lymph nodes onto a sterile wire mesh screen (approximately 3 x 3 cm2 in size) placed inside a sterile glass petri dish containing ˜10 ml of Hanks’ balanced salt solution (HBSS) (Figure 4).
- With the petri dish inclined slightly, press the lymph nodes against the screen with a syringe plunger head. Make sure to break all lymph nodes so that the cellular contents of the nodes pass through the screen while the non-cellular components of the nodes (i.e., membranes, stroma, fat) remain on the screen.
- Using the same plunger and syringe, aspirate the HBSS containing cells. Wash the screen with ˜5 ml HBSS and collect the remaining fluid into a 15 ml conical tube.
- Centrifuge at 800 x g for 10 min. Aspirate any fatty deposits at the top of the fluid with a pipette prior to discarding the fluid. Wash the cell pellet three times with HBSS. Resuspend the pellet in 1 ml of HBSS and enumerate viable cells by trypan blue dye exclusion.
7. Representative Results:
The cellular fractions of vaginal lavage fluid from >4-day inoculated mice typically consist of Candida, epithelial cells and cellular infiltrates (Figure 5). By wet-mount microscopy, Candida can be visually identified by the presence of hyphae as well as yeast (Figure 5A). Smear preparations of vaginal lavage fluid can be stained by Papanicolaou technique to examine epithelial cells and infiltrating leukocytes, of which the principal cells are neutrophils identified by the tri-nuclear lobes (Figure 5B). Very few neutrophils, if any, are detected in uninoculated mice (Figure 5C)41.
An example of vaginal fungal burden is shown in Figure 6. Vaginal lavage fluid collected at specific time points are cultured for CFU enumeration (Figure 6A). Vaginal colonization/infection with Candida persists for weeks in estrogen-treated inoculated mice (Figure 6B), while Candida fails to establish vaginal colonization in non-estrogen-treated inoculated mice (Figure 6C). Estrogen-treated uninoculated mice remain negative for Candida throughout the time (data not shown). In addition, vaginal lavages can be performed either one time on separate mice at each time point or longitudinally in the same mice under anesthesia.
The lumbar lymph nodes are the primary draining lymph nodes of the genital tract and the most relevant site to evaluate for systemic immune responses to a vaginal challenge. Note that these lymph nodes may become enlarged in inoculated mice while they normally appear quite small in uninoculated mice. Leukocyte cellular recoveries typically range from 8 × 105/uninoculated mouse to 5 ×106/inoculated mouse. In addition to lumbar lymph nodes, inguinal, popliteal and mesenteric lymph nodes can also be used.
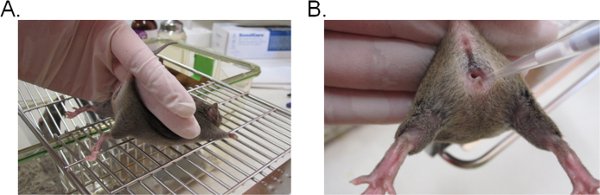
Figure 1. Vaginal inoculation with Candida. A) A mouse restrained for inoculation. The mouse is placed on a wire cage insert and held by the base of the tail, slightly upward to lift the legs and expose the vaginal opening. The hip of the mouse can be stabilized with the same hand as it attempts to resist the tail restraint. B) Introduction of the inoculum into the vaginal lumen. A pipette tip is gently inserted about 5 mm deep into the vaginal lumen. The suspension inoculum is then deposited.
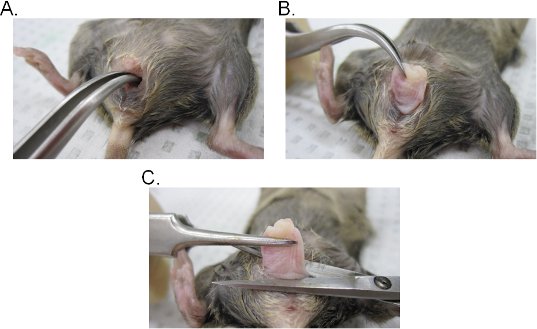
Figure 2. Vaginal tissue extraction. A-B) Extraction of the cervix. The cervix is located with curved forceps and exposed outward through the vaginal cavity. Once out of the vaginal cavity, the cervix is further pulled outward to fully expose the vagina. C) Extraction of the vagina. The vagina is excised from the vulva with scissors. Once detached, remove the cervix from the vagina.
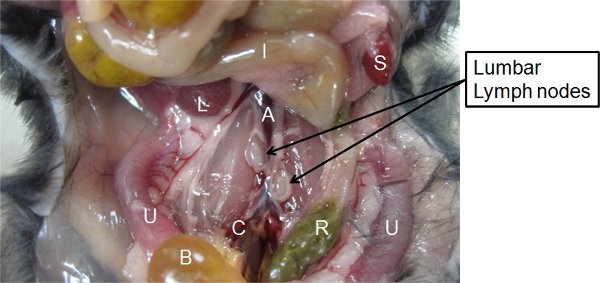
Figure 3. Identification of the lumbar lymph nodes. The location of the lumbar lymph nodes among the surrounding organs/blood vessels in the vicinity of the pelvis is indicated. A, abdominal aorta. B, urinary bladder. C, common iliac artery. I, intestines. L, liver. R, rectum. S, spleen. U, Uteri.

Figure 4. The lumbar lymph nodes placed on a wire mesh screen. The lymph nodes are pooled onto the screen placed in a petri dish with HBSS. The lymph nodes are pressed against the screen with a syringe plunger to obtain lymphoid cells in single-cell suspensions.
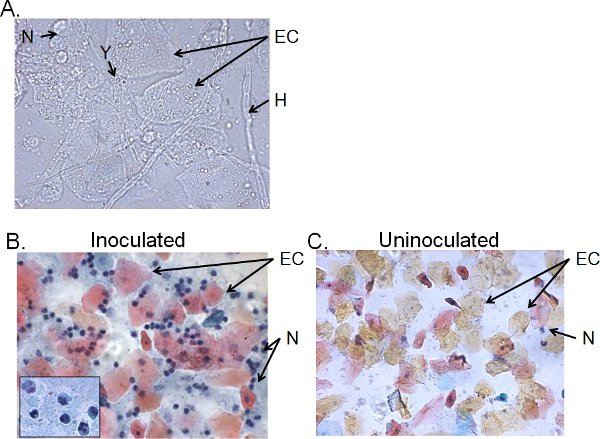
Figure 5. Cellular fractions of vaginal lavage fluid from inoculated mice. A) Wet-mount and B) Pap smear preparations of vaginal lavage samples collected 4 days post-inoculation and C) from uninoculated mice. Images are shown at 1000× (A) or 400× (B, C) magnification. The insert in B shows the nuclear morphology of vaginal neutrophils at 1000× . Candida yeast (Y) and hyphae (H), epithelial cells (EC) and neutrohils (N) are indicated.
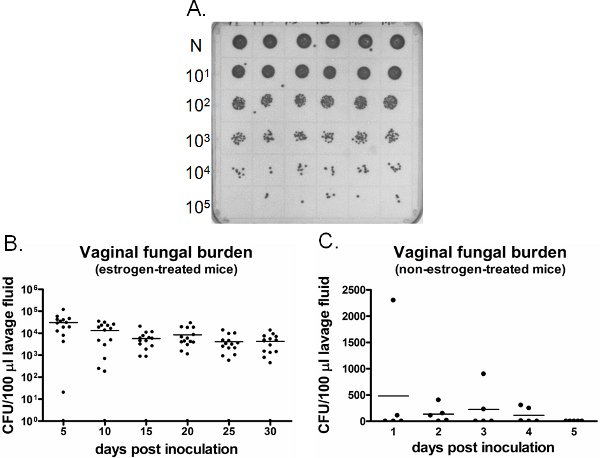
Figure 6. Detection of vaginal fungal burden. A) Representative C. albicans colonies grown on a SDA plate. Neat (N) lavage samples from six different inoculated mice (top row) were serially diluted and cultured for CFU enumeration. B) Quantification of vaginal fungal burden in estrogen-treated and C) non-estrogen-treated mice. CFU/100 μl of lavage fluid from inoculated mice was assessed on indicated time points.
