The comprehension of parasites and host cells interaction is crucial to elucidate mechanisms involved in the pathogenesis of several diseases. Although cultured human cells are less used due to limitations of cell culture compared to cell lineages, the protocol presented herein shows a robust and reproducible differentiation of human macrophages. This protocol enables the analysis of several aspects of the immune response and cell biology, from the production of inflammatory mediators up to the susceptibility of an infectious agent in human macrophages.
The first evidence that cellular differentiation is taking place is macrophage morphology (Figure 1A). On the plating day, cells are rounded and small when compared to the morphology after seven days of culture. The cellular spreading is observed when cultures are treated with macrophage colony-stimulating factor (M-CSF). In the absence of M-CSF, cell differentiation takes more time and results in an heterogenous population of macrophage-like cells (data not shown). After 7 days of differentiation, macrophages were incubated with the Alamarblue reagent for 24 hours until reading. This method allows the quantification of the cellular capacity to reduce resazurin to resorufin, thus differentiating viable from dead cells. The "ctrl" group refers to macrophages cultured for 7 days in supplemented medium, while the "dead" group refers to macrophages submitted to osmotic lysis during differentiation, which serves as a control for the technique. Once the differentiation in complete (for seven days), the macrophages derived from human monocytes remain viable and prompt to further assays that can last up to 24 hours after differentiation (Figure 1B) or few days (data not shown).
The first moments of interaction between Leishmania and a phagocyte is marked by close contact that will culminate in phagocytosis and internalization of the parasite. To understand the process of infection will help to explain the mechanisms involved in parasite killing or susceptibility to a certain pathogen. Based on the results, the first four hours of infection present the highest infection rate of L. braziliensis (both BA788 and BA3456 tested strains). After 24 hours of infection, there is a reduction in the infection rate of both strains, but we found statistical significance only for BA788 (Figure 2A, D). Considering longer periods of infection, no internalized parasites were found inside the cells after 72 hours (data not shown), suggesting that human macrophages are able to control L. braziliensis infection in vitro. The infection rate is measured by the count of 100 cells and, among those, the infected ones. This estimates the percentage of infection, which can vary due to the immune response of the cell donor, the amount of Leishmania parasites in the stationary phase and also due to the experimenter bias. Figure 2H shows representative images of a low infection rate (left) and a higher rate (right) by optical microscope.
Another data that can be assessed in cultured human macrophages is the parasite load, which is important to indicate the ability of macrophages to control the infection. It is measured by the average of internalized parasites in each cell and, after different time points, it is possible to determine whether the number of parasites has increased or reduced (Figure 2B, E). Finally, results of parasite viability further assemble information about the infection control. It is measured by the count of viable parasites in the culture after replacing RPMI to Schneider medium. Regarding L. braziliensis infection, we have already tested different time points, such as 48 h, 72 h, 96 h and 120 h to quantify viable parasites. The results indicate that 72 hours is recommended to evaluate parasite viability in these conditions (Figure 2G).
Based on our protocol, it is also possible to measure the inflammatory response against L. braziliensis infection in culture supernatant as soon as 4 hours after the infection. We were able to detect IL-6, TNF-α and LTB4 (Figure 3A-C), but IL-10 and IL-1β production was below the detection level (data not shown).
Another important aspect of the immune response against L. braziliensis is the production of reactive oxygen-derived species (ROS) by macrophages. This is one of the main mechanisms for parasite killing. The protocol presented herein show that ROS production from macrophages is significantly increased after 4 hours of infection (Figure 4A). The quantification of ROS based on corrected total cell fluorescence (CTCF) allowed the detection of almost twice the ROS production between infected and uninfected macrophages (Figure 4B).
Together, the results show that macrophage derived from humanmonocytes allows the study of several aspects of the immune response against L. braziliensis infection in vitro. Thus, this protocol enables research groups to further explore the role of human macrophages in leishmaniasis, minimizing the bias of lineage or murine cell models.

Figure 1. Cell morphology during human macrophage differentiation in vitro. (A) Representative images of the morphology of adherent cells at first day of culture (left) and after seven days of differentiation. (B) Cell viability after culture for seven days for differentiation. Ctrl = macrophages in culture with supplemented medium; Dead = macrophages subjected to osmotic lysis; Objective 60x; n = 3. Please click here to view a larger version of this figure.
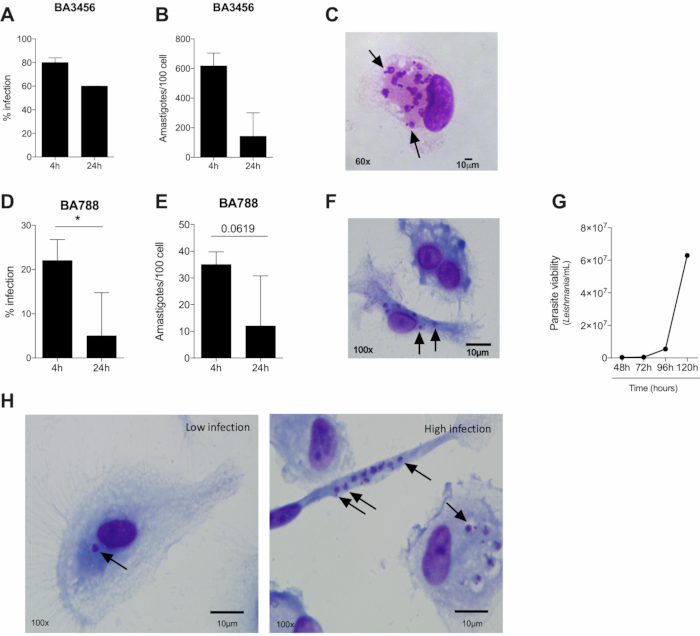
Figure 2. Several parameters of infection caused by two strains of Leishmania braziliensis can be assessed by optical microscopy. (A, D) Infection rate (B, E), parasite load (four and 24 hours of infection) and (C, F) representative images of macrophages infected by L. braziliensis from BA5456 (A-C) or BA788 (D-E) strains (four hours of infection). (G) Macrophage-derived viable L. braziliensis after four hours of infection with BA788 strain. (H) Representative images of human macrophages infected by L. braziliensis (BA788 strain) showing low (left) and high infection rate (right). Arrows = intracelular amastigotes; Scale bar 10 µm; *p < 0.05; (A,B) n = 3; (D,E) n = 6; (G) n = 6. Please click here to view a larger version of this figure.
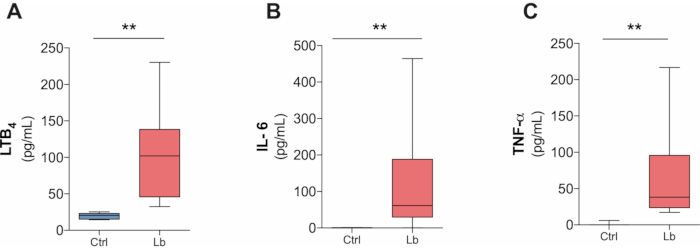
Figure 3. L. braziliensis-induced production of inflammatory mediators by human macrophages in four hours after infection. (A) LTB4, (B) IL-6 and (C) TNF-α production in culture supernatant after four hours infection by L. braziliensis (BA788 strain) measured by ELISA. **p < 0.01; n = 5. Please click here to view a larger version of this figure.
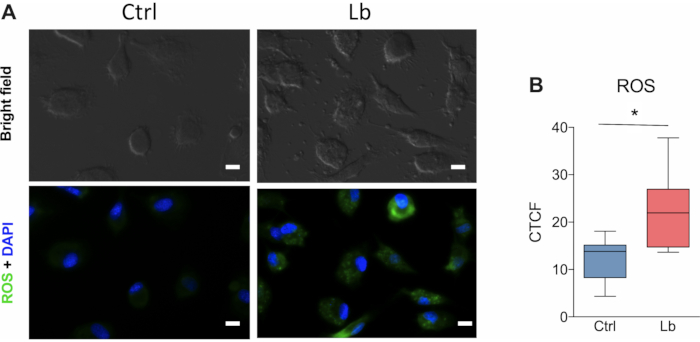
Figure 4. ROS production by human macrophages after in vitro infection with L. braziliensis. (A) Representative images of ROS fluorescent labelling in cultured human macrophages after four hours of infection by L. braziliensis (BA788 strain). (B) Quantification of ROS production based on the corrected total cell fluorescence (CTCF) using Image J. Green = ROS; Blue = nucleus; Scale bar 10 µm; * p < 0.05; n =6. Please click here to view a larger version of this figure.
