An overview of a typical maternal intranasal immunization study design is depicted in Figure 1, incorporating the immunizations, breeding, kindling, lactation, and antibody transfer. Though not illustrated, blood should be collected prior to the initial immunization for baseline measurements and throughout the remainder of the study at regular intervals. Blood is easily obtained via the central ear artery with mild sedation and a topical analgesic agent (e.g., lidocaine 2.5% and prilocaine 2.5% cream). The presence of antigen-specific IgG levels can be measured in these samples. Female rabbits are immunized via the intranasal route, as described in the protocol and demonstrated in the video. Depending on the study, the vaccine may require a boost or may need to be given through an additional route (e.g., intramuscular or subcutaneous). Following study initiation, rabbits are bred; we prefer to purchase proven breeders from vendors to use to ensure a higher pregnancy rate for these studies. Depending on the immunization timeline, rabbits may receive additional immunizations throughout pregnancy. Antigen-specific IgG is transferred transplacentally to the kits, and at approximately 30-32 days post-breeding, pregnant does will kindle. We recommend limiting handling of kits for the first several days to minimize rejection from the does. Blood samples can be collected from the kits to assess antigen-specific IgG levels that were transferred transplacentally (Figure 3). In addition to a wide variety of nutrients, kits receive IgA from the lactating doe while nursing. Kits are typically weaned at 4-8 weeks, but prior to weaning, milk can easily be collected from lactating does, as demonstrated in the video. The collected milk samples can then be processed for the detection of total and antigen-specific IgA levels (Figure 4). Depending on the study, vaccines (+/- boosts) can be administered to the kits, and serial blood samples can be collected from the kits at a very early age using the lateral saphenous vein.
For maternal studies, determining pregnancy as early as possible is helpful for the study design and for ensuring the doe does not need to be rebred. Progesterone measurements can be used as a means to detect pregnancy. As shown in Figure 2, elevated progesterone levels can be detected in pregnant rabbits compared to non-pregnant rabbits even after matings by a buck were confirmed for all does. There are additional methods for pregnancy detection, including manual palpation, ultrasound, and radiographs; however, these require well-trained personal and proper equipment.
Antigen-specific IgG that was transferred transplacentally while in utero can be measured in the serum of kits. Blood can be collected from a small number of kits at or near the time of birth to assess early antigen-specific antibody levels, but serial blood collection is technically much easier as the kits age and increase is size. As depicted in Figure 3, serum levels of antigen-specific IgG in the kits can be measured by ELISA and compared to the maternal levels. Maternally transferred IgG levels tend to be higher at birth and decrease over time.
As a type of mucosal sample, milk can be collected and processed to measure total or antigen-specific antibody levels. As shown in Figure 4, IgA makes up a significant portion of the total antibody levels within the breast milk that is being transferred to the kits via lactation. Our results demonstrate that breast milk IgA produces a slightly higher ELISA signal (relative light units, RLU) when compared to IgG, and both IgA and IgG produce a signal that is much higher than the signal for IgM. These results are in agreement with results from others that suggest that rabbit milk contains around 4.5 mg/mL IgA, 2.4 mg/mL IgG, and 0.1 mg/mL IgM26,27.
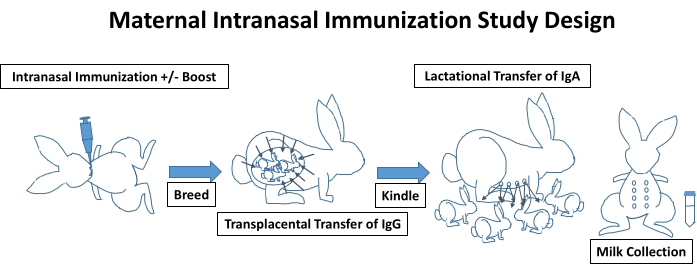
Figure 1. Sample timeline for a maternal intranasal immunization study design in a rabbit (Oryctolagus cuniculus) model. Female rabbits are immunized via the intranasal route, as described in the protocol and demonstrated in the video. Depending on the study, the vaccine may require a boost or may need to be given through an additional route (e.g. intramuscular or subcutaneous). Rabbits are then bred. Antigen-specific IgG is transferred transplacentally to the kits, and at approximately 30-32 days post-breeding, pregnant does will kindle. IgA is passed to the kits from the lactating doe while nursing. Prior to weaning, milk can easily be collected from the lactating does to assess total and antigen-specific IgA levels. Please click here to view a larger version of this figure.
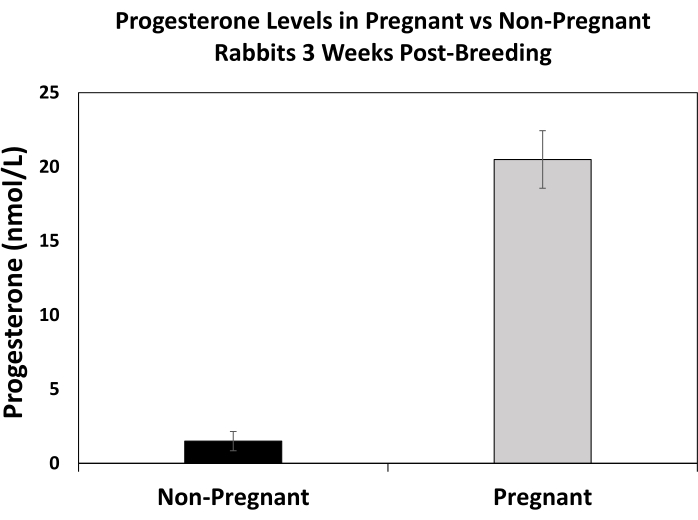
Figure 2. Progesterone levels in pregnant and non-pregnant rabbits at 3 weeks post-breeding. Blood was collected from rabbits at 3 weeks post-breeding. Rabbits were confirmed either pregnant or non-pregnant based on ability to kindle a litter at 30-32 days post-breeding. Serum progesterone levels were measured through the Michigan State University Veterinary Diagnostic Laboratory using a chemiluminescent immunoassay (CLIA) with an immunoassay system (e.g., Siemens Healthineers IMMULITE 2000). Error bars represent standard error of the mean, and the sample size consisted of 4-6 rabbits per group. Please click here to view a larger version of this figure.
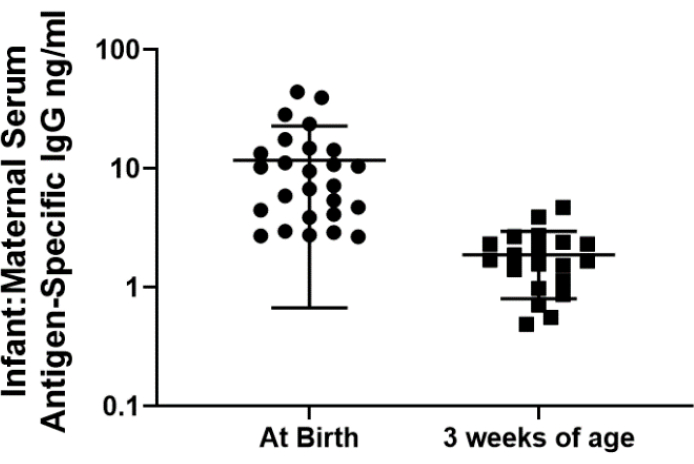
Figure 3. Antigen-specific IgG levels in kits (relative to maternal levels) at birth and at 3 weeks of age following a series of maternal immunizations. Blood was collected from the does and kits soon after kindling and at 3 weeks of age. Antigen-specific IgG levels within the serum were detected using a fluorescent ELISA as previously described28. Antigen-specific IgG is plotted as a ratio of levels detected in the kit serum and maternal serum. Please click here to view a larger version of this figure.
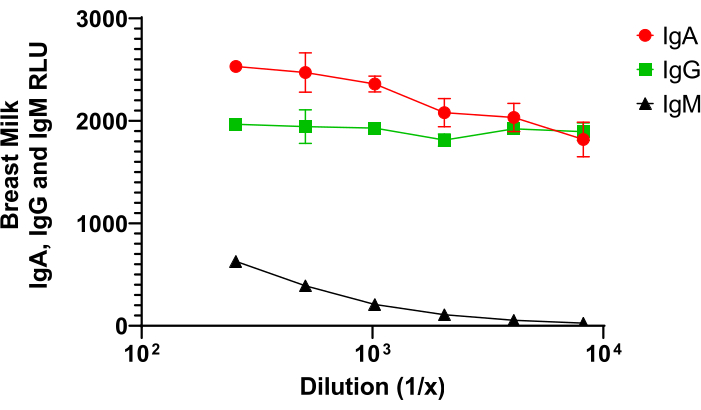
Figure 4. A comparison of IgA, IgM, and IgG levels in rabbit milk. Rabbit milk was collected as described and demonstrated in the video. Milk was processed by a long centrifugation (13,000 x g for 4.5 hours at 4 °C), and the clear middle layer was isolated following processing. Total IgA, IgG, and IgM levels were measured in this clear layer by fluorescent ELISA as previously described28, except that plates were coated with polyclonal anti-IgA, anti-IgG, or anti-IgM to detect total rabbit IgA, IgG, or IgM, respectively. Please click here to view a larger version of this figure.
