Analysis of C. elegans locomotor activity in the liquid media could easily capture an integrated phenotype of mitochondrial disease worm models that may not be easily quantifiable on solid media. ZebraLab was used to quantify locomotor activity of the well-established mitochondrial complex I disease gas-1(fc21) strain relative to WTworms in liquid media at the L4 larval stage. The activity of 5 worms in a single liquid drop was recorded over 1 min, with a total of 19 videos (technical replicates) recorded for each strain, resulting in total the analysis of 95 worms per strain. Four biological replicate experiments were obtained per strain. Worm activity is displayed as pixel change (Figure 3A), and as percent of control (POC) when normalized to N2 Bristol WT control (Figure 3B). The gas-1(fc21) worms (62% ± 16% pixel change, mean ± SD, n = 19) had a significant 38% decrease (p < 0.001, t-test) in their locomotor activity at L4 stage as compared to WT worms (100% ± 11.35%, mean ± SD, n = 95 worms per condition in 19 technical replicates over 4 biological replicates).
WormScan analysis was also performed to quantify the locomotor activity of L4 stage gas-1(fc21) and WT worms in liquid media. Data was collected for three biological replicate experiments, where each biological replicate plate was evaluated by two sequential images scanned using a standard flatbed scanner. Worm activity of the differential images was compared as pixel change and normalized to concurrent N2 Bristol WT control. Similarly, as was seen by the Zebrafish behavior screening method, WormScan based analysis demonstrated that the gas-1(fc21) worms (65.9 ± 6.1, mean ± SD, n = 13 wells) had a significant decrease in locomotor activity by 34% (p < 0.001, t-test) compared to N2 Bristol wild-type worms (100% ± 4.8%, mean ± SEM, n = 12 wells) (Figure 3C). Analysis using WormScan on day 1 adult gas-1(fc21) worms (50.1% ± 10.7%, mean ± SD, n = 7 wells) demonstrated a decrease in locomotor activity by 49% (p < 0.001, t-test) compared to WT worms (100% ± 16.2%, mean ± SD, n = 6 wells) (Figure 3D).
Table 1: Comparative overview of experimental assays available to evaluate C. elegans neuromuscular activity. A detailed overview is provided of a wide array of 16 different experimental techniques that can be used to quantify worm neuromuscular activity on the phenotypic outcomes of thrashing, locomotion, pharyngeal pumping, and/or chemotaxis in C. elegans. Read format, methodology, experimental throughput capacity, software and/or equipment requirements, as well as advantages and limitations of each assay are detailed. References and relevant websites for each assay and software tool are also provided. The throughput capacity of each assay is described as low, medium, or high, as based on the experimental complexity, the use of single or multi-well plates, and/or experimenter time needed to complete the experimental setting and data analyses. * Indicates that the methodologies can also be used for evaluation of locomotion. Please click here to download this Table.

Figure 1: C. elegans locomotor activity analysis using ZebraLab software. (A,B) Experimental protocol for worm video recordings. Five worms were introduced per drop (20 µL) of S. basal solution, with four drops placed on a single glass slide under a stereomicroscope. Each drop of 5 worms represented a technical replicate experiment and was recorded for 1 min in a separate movie using a charged-coupled device (CCD) camera. (C–F) Experimental settings in ZebraLab as adapted to evaluation of locomotor activity in C. elegans. (C) Selection of Quantization with AVI files to quantify worm locomotor activity of each recorded video. (D) Protocol parameter settings, with 1 min selected as experiment duration. (E) Build area to select the area of interest. The area was selected and built around 1 drop of solution in which 5 worms were placed. (F) Detection was determined based on gray-scale thresholding to detect the whole body of each worm (red). In the threshold section, burst and freezing values were selected to analyze worm activity as pixel changes. Please click here to view a larger version of this figure.
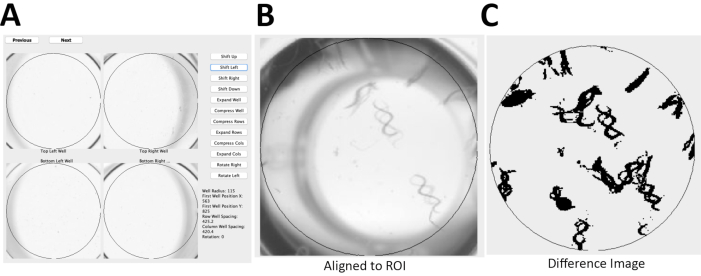
Figure 2: C. elegans locomotor activity analysis using WormScan methodology. (A) Using an Epson v800 flatbed scanner, two immediately sequential scans of a 96-well plate were captured with a resolution of 1,200 dots/in and 16-bit grayscale to produce jpeg images. (B) These two sequential images of a 96-well plate were then aligned to a reference region of interest (ROI), of WT worms. (C) Image analysis is based on a difference image score calculated for each ROI with 15 worms/well for N2 Bristol. The difference image was normalized and reported as percentage of control (POC). Please click here to view a larger version of this figure.
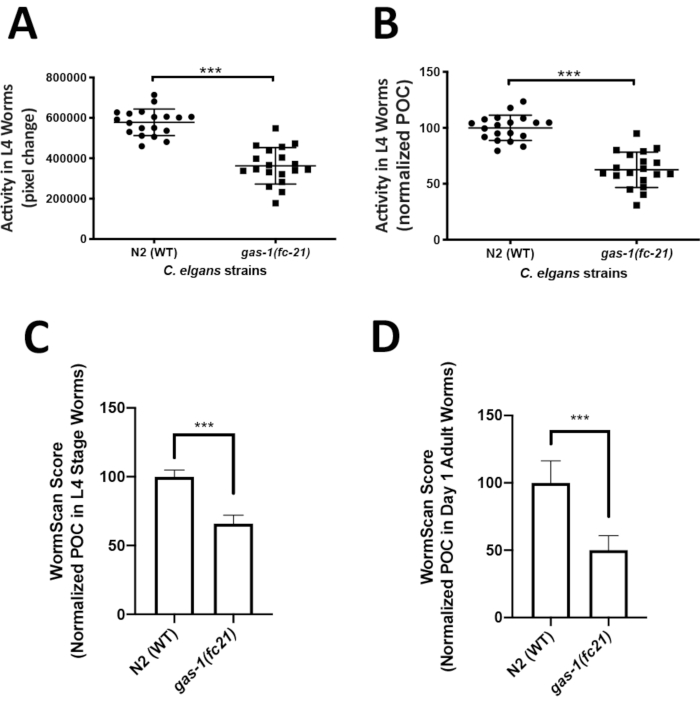
Figure 3: Comparative analysis of locomotor activity by ZebraLab and WormScan software assays in gas-1(fc21) mitochondrial disease worms relative to N2 Bristol wild-type worms. (A,B) WT and gas-1(fc21) worm activity in liquid drops (5 worms/drop) was video recorded for 1 min and quantified as (A) pixels change or (B) percentage of wild-type control using the ZebraLab software. Overall, ZebraLab-based worm activity analysis demonstrated a significant decrease by 38% in gas-1(fc21) L4 stage worms as compared to wild-type controls (*** p < 0.001). The graph displays mean ± SD of all data, where each dot conveys the overall activity of five worms per S. basal drop. Each drop represents a technical replicate, with a total of four biological replicates studied per condition. A total of 19 videos were recorded (one video for each drop of 5 worms), across a total of 95 individual worms studied per condition. Statistical analysis was performed using the student t-test in Prism -GraphPad v6. (C) WT and gas-1(fc21) worms at L4 stage were analyzed by flatbed scanning to produce two sequential images that were analyzed in WormScan software to yield a difference image. Three biological replicate experiments were performed with 15 worms per well in a 96 well-plate. The activity of WT worms was used as the baseline to normalize percentage of control (POC). gas-1(fc21) activity was decreased by 34% as compared to wild-type control (*** p < 0.001). Bar graphs convey mean and standard deviation across three biological replicate experiments. (D) N2 and gas-1(fc21)worms at adult day 1 stage were analyzed similarly as detailed for panel C. gas-1(fc21) activity in day 1 adults was decreased by 49.1% relative to wild-type control worms (*** p < 0.001). Bar graphs convey mean and standard deviation of pixel changes in one biological replicate comparing N2 (n = 6 wells of 15 worms/well) and gas-1(fc21) (n = 7 wells of 15 worms/well). Please click here to view a larger version of this figure.
