Figures 1 and 2 display the experimental setup and equipment used to record protein activity in the course of mechanical stimulation of the lipid bilayer membrane. To minimize electrical noise into our measurements, the workstation is placed within a lab-made Faraday cage, grounded to a ground connection on the AxoPatch 200 B Amplifier.
Formation of a stable insulating lipid bilayer is a key step in this study. In this arrangement, a lipid monolayer assembles at the oil/water interface of the aqueous droplets immersed in a bath of an organic solvent. When droplets are placed in contact, excess oil is eliminated, and the opposing lipid monolayers thin to a two-molecule thick lipid bilayer. The most common technique used in bilayer characterization is voltage-clamp. With voltage-clamp, the voltage across the bilayer is maintained at a constant value while the current is measured. Figure 4 portrays a typical real-time current recording of the initial bilayer formation. Knowing the specific capacitance (~0.6 μF/cm2)5 of the DPhPC lipid bilayer, the area of the formed bilayer could be calculated. The bilayer area could be controlled by changing the position of the droplets (Figure 4A). Using the piezoelectric actuator, different types of waveforms (sinusoidal, square, triangular, etc.) at different frequencies, amplitudes, and duty cycles could be applied to the droplets to horizontally and axially oscillate them and thus, bilayer tension and area could be altered (Figure 4B).
When the DIB is mechanically stimulated, while maintaining a constant DC potential across the membrane, a low-threshold (gain-of-function) V23T mutant of MscL generates reliable activities including mainly sub-conductive states and occasionally full opening events (Figure 5). These events are identical to those recorded using the patch-clamp technique from intact inner E. coli membranes and liposomes reconstituted with the purified V23T MscL. The results in Figure 5 prove that gating occurs in response to an increase in tension, since all current spikes are observed at peak compression. At peak compression, the relative areal expansion of the droplets is maximal and therefore, tension at the interface is maximal.
Alamethicin, a voltage-gated ion channel and one of the most studied peptides, increases the membrane permeability when a DC voltage is applied across the membrane36. The ability of the lipid bilayer interface to host transmembrane proteins and peptides is also tested by performing voltage-gating current recordings using alamethicin peptide. Alamethicin is mixed with the phospholipid solution to a final concentration of 100 ng/ml. Figure 6 shows the current measurements under voltage clamp (+115 mV). The droplets in this experiment are pulled apart in order to achieve small bilayer interface and thus higher resistance and smaller capacitance. The gating behavior of the Alamethicin peptide is shown through the discrete steps of current (Figure 6). The histogram on the right side of the plot shows the changes in conductance from the base level (0.0962 nS), which is basically the first conductance level of the channel itself.

Figure 1: A schematic describing the main parts and dimensions of the oil reservoir. The oil reservoir is manufactured at the machine shop at Virginia Tech. It consists of a machined cylindrical acrylic tube glued to the surface of an acrylic sheet. The dimensions and design can be modified to accommodate different applications or more than two micropipettes. Please click here to view a larger version of this figure.
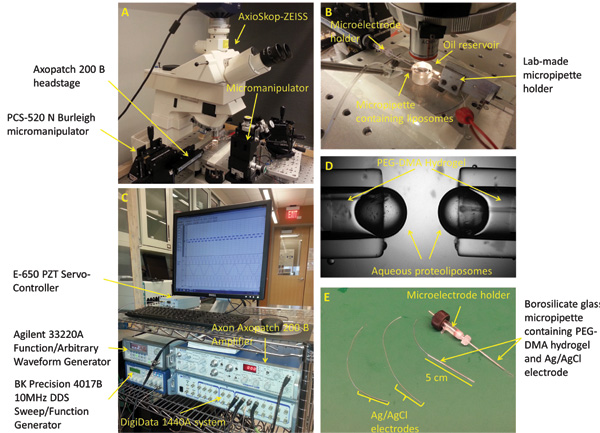
Figure 2: Experimental setup and micropipettes preparation. (A) The standard workstation for forming, mechanically stimulating, and characterizing the interface bilayers includes a microscope, 3-axis manipulators, a digital camera, piezoelectric oscillator, vibration isolation table, and a Faraday cage (not shown). (B) The experimental setup consists of two opposing PEG-DMA hydrogel filled micropipettes horizontally positioned within a bath of Hexadecane oil. Each of the micropipettes contains an Ag/AgCl electrode to provide electrical connection. A third micropipette filled with proteoliposome solution is used to form the droplets at the tip of the other micropipettes. (C) The DIB current response could be measured using a combination of the patch amplifier and the low-noise data acquisition system. (D) A closed up picture showing the aqueous droplets formed at the tip of the micropipettes. (E) Ag/AgCl electrodes are made by dipping the tip of two 250 μm silver wires in bleach. The electrodes are then fed through two borosilicate glass capillaries filled with PEG-DMA hydrogel, which is cured with UV light to solidify. A straight microelectrode holder with male connector is used to connect one of the micropipettes to the headstage of the patch amplifier.
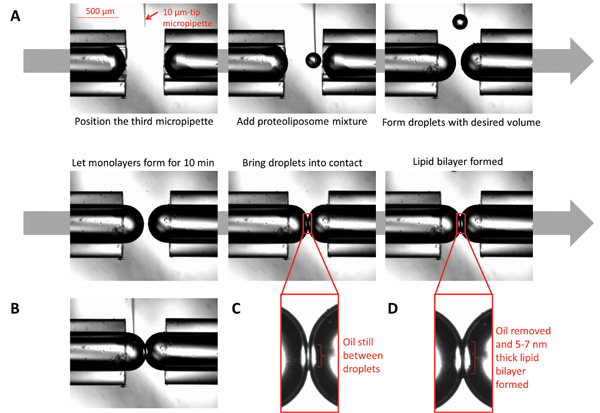
Figure 3: Images illustrating the formation of droplet interface bilayers. (A) A 10 μm micropipette filled with proteoliposomes is positioned under the microscope in proximity to the micropipette tips. Using a syringe connected to the micropipette, dispense small volumes of the proteoliposomes to form spherical droplets to desired volume. Let the monolayer form by allowing the droplets to sit for ten min. Bring the droplets into contact; the bilayer will form after all oil at the interface is eliminated. (B) While the bilayer is formed, the chemical composition at both sides of the interface could be controlled by injecting desired chemicals using a micro-sized micropipette. (C) The droplets at the moment of first contact. (D) The droplets when the lipid bilayer is formed.
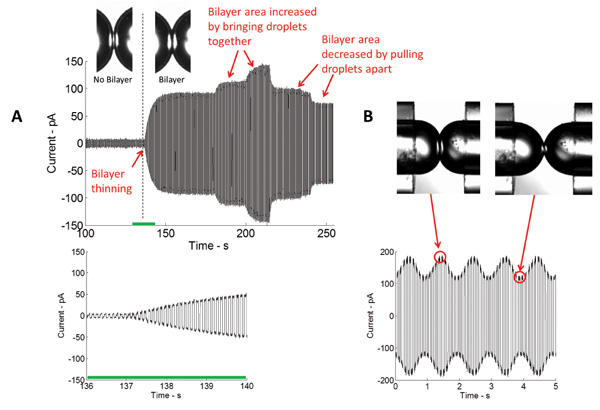
Figure 4: Real-time measurements show both the initial thinning and subsequent expansion of the interface. (A) Current measured in the course of bilayer formation through the application of a triangular electrical potential. The magnitude of the measured current is directly proportional to the capacitance, and thus the area of the bilayer interface. The closer the droplets are brought together, the bigger the area of the interface and vice versa. (B) Upon application of mechanical excitation, the area of the bilayer interface increases and decreases at the same frequency as the stimulating signal.
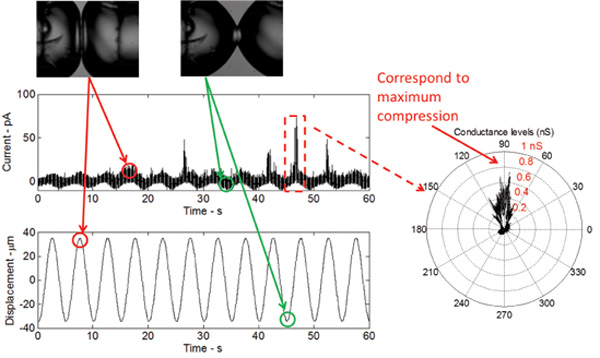
Figure 5: Real-time measurements show the response of the bilayer to mechanical excitation as well as the gating of the V23T mutant of MscL. The shape of the current response is sinusoidal, which relates to a sinusoidal change in bilayer capacitance as a result of the bilayer area change. The current spikes, occurring at the peak of each cycle, indicate sub-conductance gating of the V23T mutant. A polar plot further indicates that gating occurs at peak compression, which reflects an increase in tension at the bilayer interface.
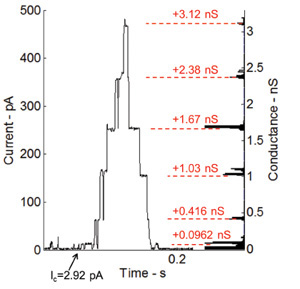
Figure 6: Current measurements under voltage clamp and corresponding histogram of conductance levels for gating activity of incorporated Alamethicin channels. The gating behavior of the Alamethicin peptide is shown through the discrete step-wise increase in current. The conductance levels match very well with previous measurements performed by our research group at Virginia Tech7.
