Example: When foraging, C. elegans transitions from forward to reverse motion, often performing a reorientation (omega turn) before returning to the forward motion state. Quantifying this transition is important in understanding the foraging patterns of movement and also in the worm’s motor control. The power to reveal subtle details of locomotion behavior can be seen using our tracker device.
As an example we look at the forward to reverse and reverse to forward transition by capturing high resolution images of worms crawling freely on the agar plate for ~30 min. runs. The eigenmode analysis (Figure 3) allows us to measure the phase of the undulatory movements of the worm. The phase measurement in turn allows us to calculate the velocity of the worm and the forward to reverse transition (Figure 4). By plotting the joint probability of the worm’s velocity and phase, we can see that the transition between forward and reverse happen probabilistically with a preferential phase (Figure 5a). If we separate forward to reverse and reverse to forward transitions and look at the conditional probability of the phase of the worm when it enters and exits a reversal event, we see that they have distinct phase distributions (Figure 5b,c).

Figure 1. Tracking microscope. The sample of a worm crawling on an agar plate remains stationary while the imaging system moves to keep the worm in the center of view. The imaging system is built on a 2D translation stage driven by stepper motors (3). A CCD camera (Basler A601f) and lens (25mm focal length) images the worm from below (1) and a fiber light (Edmunds optics; model) illuminates it from above (2). A homemade stepper motor controller utilizing a SimpleStep controller board (SimpleStep; SSXYZ) controls the movement of the imaging system. A custom LabVIEW program (National Instruments, Austin, TX, USA) acquires and processes images of the worms, while simultaneously communicating with the motor controller to keep the worm in the center of the field.
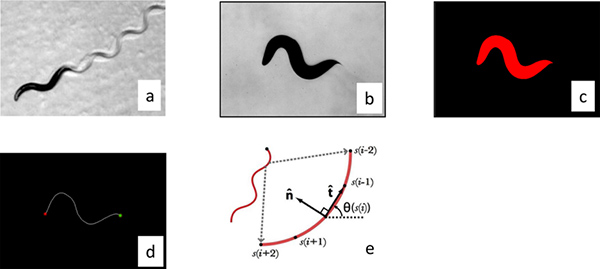
Figure 2. Image processing. (a) Worms move by changing their curvature in time and so we processed the worm images by parameterizing their center line. The images were made binary by thesholding the raw gray scale image (b), and individual worms were identified by filtering objects by size (c). The distance between the center of the mass of the worm and the center of the image was calculated and then the stage was moved the appropriate distances to re-center the worm. These correction distances were used to calculate the trajectory of the worm. The curvature of the worm perimeter was used to identify the tail (maximum curvature) and head (secondary maximum). (d) The images were then skeletonized to locate the center line. (e) The resulting curve was then then interpolated into 101 segments and the calculated angle between segments was used to parameterize the curve. Click here to view larger figure.
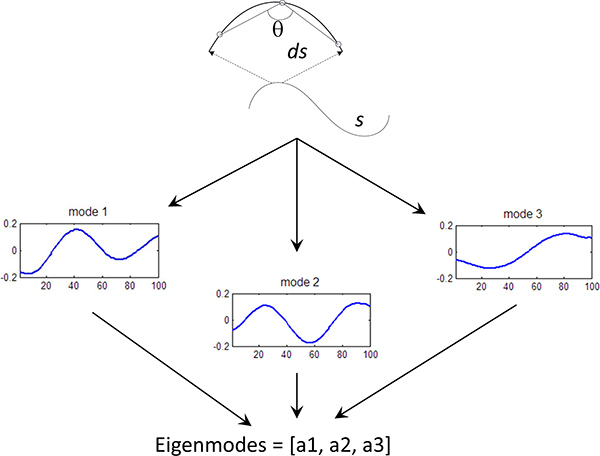
Figure 3. Eigenmode calculation. Fundamental curves of the worm shape have been calculated using Principle Component Analysis5. Each worm curve is represented by a linear sum of the eigenvectors (or eigenworms), where the amplitude of each components is known as the eigenmode (mode). We can quantify the behavior of the worm by measuring the first 3 eigenmodes of the worm shape over time.
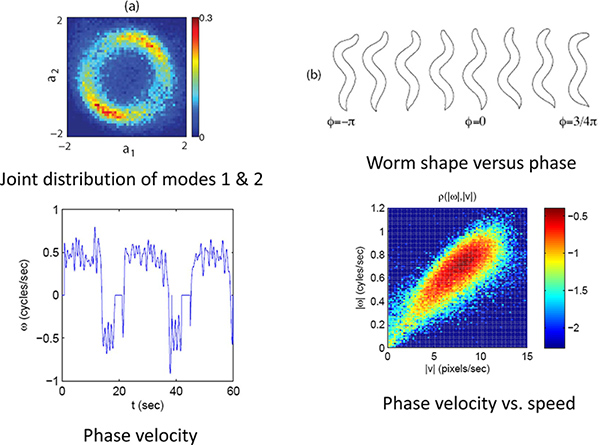
Figure 4. Phase and velocity. (a) The joint distribution of the first two modes for a limit cycle. (b) The phase or position on this limit cycle correlates with the phase of the worm’s undulatory position. (c) Using the phase velocity the worms forward and backward movements can be quantified. (d) The speed and direction along this limit cycles indicates the worm’s velocity. Click here to view larger figure.
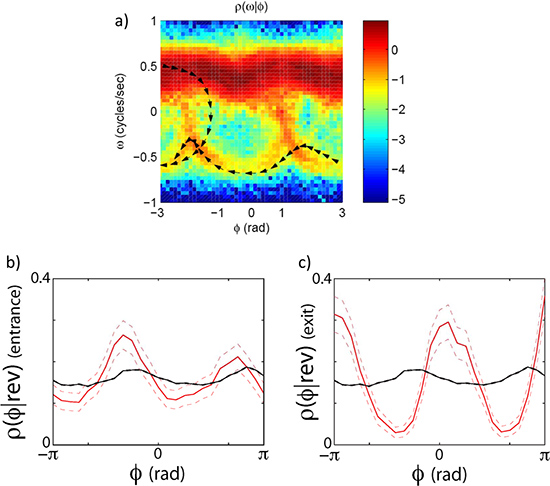
Figure 5. Phase dependent velocity switching. The transitions between forward and reverse states do not happen randomly along the oscillatory cycle. They (but still stochastically) happen at specific phases. (a) The joint distribution of the phase velocity and speed clearly indicates that worms leave and enter the forward state preferentially at specific phases. (b) shows a plot of the phase conditioned on the entrance to a reversal event. (c) shows the phase conditioned on the exit of a reversal event. The black line is the distribution of phase angles without conditioning on an exit or entrance.
